



Plain-nosed bats (family Vespertilionidae) as a possible reservoir of lyssaviruses and coronaviruses in Western Siberia and the south of European Russia
- Authors: Ohlopkova O.V.1,2, Kononova Y.V.2, Tyumentseva M.A.1, Tyumentsev A.I.1, Shestopalov A.M.2, Akimkin V.G.1
-
Affiliations:
- Central Research Institute of Epidemiology, Rospotrebnadzor
- Virology Research Institute of the Federal Research Center for Fundamental and Translational Medicine
- Issue: Vol 69, No 5 (2024)
- Pages: 415-428
- Section: REVIEWS
- URL: https://virusjour.crie.ru/jour/article/view/16685
- DOI: https://doi.org/10.36233/0507-4088-267
- EDN: https://elibrary.ru/vpenkk
- ID: 16685

Cite item

Abstract
The review presents current data on the chiropterofauna inhabiting Western Siberia and the south of the European part of Russia. A general description of the genus of lyssaviruses and the family of coronaviruses is given. The potential for virus carriage in relation to lyssaviruses and coronaviruses in bat populations of two geographically distant regions is considered.
Full Text
Introduction
Occasional outbreaks of infectious diseases caused by viruses necessitate research aimed at finding foci of such diseases and assessing the risk of new viral epidemics. Thus, the study of the SARS-CoV-2 virus, the COVID-19 pathogen, and the routes of its transmission to humans has revealed the important role of bats as natural reservoirs of potentially dangerous viruses. Furthermore, bats are the source of many species of lyssaviruses, the scale of their spread, high pathogenicity and almost absolute lethality determine their relevance and primary importance for the study of lyssaviruses.
This literature review focuses on the fauna of bats and the potential for virus carriage in certain members of this group in Western Siberia and southern European Russia. These geographic areas are of particular interest for studying the circulation of lyssaviruses and coronaviruses for several reasons. First, both regions are characterized by significant biodiversity and the presence of different ecosystems and biotopes. Second, the geographical distance of these areas makes it possible to conduct a comparative analysis of viruses circulating in different climatic and ecological conditions. It is also worth mentioning separately the proximity of the Siberian Federal District to China, where, due to various reasons, foci of new viral infections may arise. Thirdly, the analyzed regions are characterized by high population density and active anthropogenic activity, which creates conditions for interaction between humans and wild animals, and potentially increases the risk of interspecies transmission of viruses.
Currently, RNA- and DNA-containing vertebrate viruses belonging to 35 families have been isolated or detected by molecular genetic methods in representatives of the order Chiroptera [1]. The frequency of viral RNA/DNA occurrence in animal organisms varies among species of bats and depends on both the virus and its host (vector) and geographic distribution [1, 2]. Most of the viruses identified in bats are not associated with human diseases; however, these animals are known to carry and transmit viruses pathogenic to humans (rabies virus, Nipah virus, Marburg virus, etc.). In particular, 4 lethal cases of lyssavirus infection after contact with plain-nosed bats were reported in Russia during the years 1977–2007 (Lugansk city – 2 cases; Belgorod region – 1 case; Primorsky Krai – 1 case) as well as 3 new cases in the period of 2019–2021 in the territories of the Far East [3]. Thus, when assessing the epidemiological significance of bats, a combination of factors should be taken into account: species diversity, abundance, ecological conditions and behavior, associations with known viruses.
According to the information given in the international database Higher Taxonomy (https://www.mammaldiversity.org/taxa.html), 52 species of bats are currently registered on the territory of Russia. The Vespertilionidae family constitutes the major part of the chiropterofauna of Russia and includes 18 species of mouse-eared bats (Myotis), 5 species of long-eared bats (Plecotus) and Pipistrellus each, 4 species of noctule bats (Nyctalus) and house bats (Eptesicus), 3 species of broad-eared bats (Barbastella), and 2 species each of pipistrelles (Hypsugo), vesper bats (Murina) and parti-colored bats (Vespertilio). Plain-nosed bats are found throughout Russia, from parts of the tundra in the north [4] to subtropical areas in the south [5], from the Baltic coast in the west [6] to the Far East [7–9]. The above-mentioned causes considerable research interest in this particular family of bats, as the most actively contacting with the human population, through settlement near human dwellings, hibernation in attics and basements.
Bats of other families are represented by 4 species of horseshoe bats (Rhinolophus), two species of longwings (Miniopterus) and one species of free-tailed bats (Tadarida). Representatives of 4 species of horseshoe bats, the common longwing (M. schreibersii) and the European free-tailed bat (T. teniotis) are found in some areas of the south of the European part of Russia – in the Caucasus and the Black Sea region [5, 10], the range of the eastern longwing (M. fuliginosus) in Russia is limited to the south of Primorsky Krai [7].
The species diversity of Chiroptera depends on a combination of factors, the main of which are the presence of favorable environmental conditions and availability of food resources. As a consequence, the greatest number of species is characteristic of equatorial, tropical and subtropical latitudes, with a gradual decrease to the north in the Northern Hemisphere and to the south in the Southern Hemisphere [11]. The Rostov and Novosibirsk regions, studied as risk territories in the presented analysis of scientific literature, are located within the temperate climatic belt of Northern Eurasia, but differ significantly in average annual temperatures, landscape diversity, and the degree of anthropogenic transformation of natural biocenoses. The study of the virus circulating in these two distant regions can provide insight into the nature of genetic diversity and the presumed limits of distribution of individual species or genetic variants of viruses. The Chiroptera fauna and viruses of these regions together form a single virome landscape represented on the Eurasian continent, in which viruses circulating among representatives of the family of plain-nosed bats occupy a significant place.
The aim of this study is to analyze the potential of virus carriage with respect to lyssaviruses and coronaviruses in populations of plain-nosed bats in two remote regions of the Russian Federation. The information obtained on the relationship between the Chiroptera fauna and species susceptibility to viruses potentially dangerous to humans can be used to predict epidemiologic risks in the system of monitoring of some virus infections associated with bats.
Characterization of lyssaviruses and coronaviruses circulating among members of the family Vespertilionidae
Lyssaviruses (Rhabdoviridae: Lyssavirus). Lyssaviruses are one of 13 genera of the Rhabdoviridae family. The genome of lyssaviruses is represented by single-stranded linear negative stranded RNA with a length of about 11,9–12,3 thousand nucleotides. The virions consist of two structural units. The length of virions ranges from 130 to 380 nm. The genetic structure of lyssaviruses is conserved and is represented by 5 structural protein genes (3’ – N (nucleoprotein) – P (phosphoprotein, RNA replication cofactor) – M (matrix protein) – G (surface glycoprotein) – L (viral RNA polymerases) – 5’) [4].
Lyssaviruses are neurotropic pathogens that cause acute progressive encephalomyelitis. Sensitivity to all members of all mammalian genera is due to the commonality of pathogenesis - interaction with the nerve terminal nicotinic acetylcholine receptor, which is a universal mediator of nerve impulse transmission in synaptic connection [5]. Transmission of infection occurs through epizootic chains of vector type, arising from traumatizing contacts with a sick source animal. As a result of infection, the pathogen penetrates into motor or sensory neurons and spreads centripetally through the central nervous system using retrograde axonal transport along neuronal connections [11, 12].
In the natural environment, lyssaviruses are associated with specific reservoir animals, predominantly raptors and bats, within parasitic systems of two archetypes – terrestrial and air-terrestrial. The main hosts of lyssaviruses in the phylogenetic relation of their natural-historical origin are the representatives of the bat species. Carnivores enable the circulation of rabies virus (RABV) only. The following lyssaviruses are associated with bats of temperate climatic belt of Eurasia: European bat 1 lyssavirus – EBLV-1 (Lyssavirus hamburg according to the current ICTV classification), European bat 2 lyssavirus – EBLV-2 (L. helsinki), Aravanvan lyssavirus – EBLV-2 (L. helsinki), Aravan lyssavirus – ARALV (L. aravan), Khujand lyssavirus – KHULV (L. khujand), West Caucasian bat lyssavirus – WCBLV (L. caucasicus), Irkut lyssavirus – IRKLV (L. irkut), Bokelelo lyssavirus – IRKLV (L. irkut), Bokeloh bat lyssavirus – BBLV (L. bokeloh), Lleida bat lyssavirus – LLEBLV (L. lleida) and Kotalahti bat lyssavirus – KBLV (L. kotalahti). As a rule, the distribution of particular lyssaviruses is species-specific, but does not exclude their transmission to other species in common colonies or to occasional hosts such as domestic animals or humans [12].
Coronaviruses (Coronaviridae: Alphacoronavirus, Betacoronavirus). The Coronaviridae family is a monophyletic cluster in the Nidovirales order. They are enveloped plus-RNA viruses that affect vertebrates of 3 classes: mammals (coronaviruses and toroviruses), birds (coronaviruses) and fish (bafiniviruses). Virions can be spherical, 120–160 nm in diameter (Coronavirinae), rod-shaped, 170–200 × 75–88 nm (Bafinivirus) or a mixture of the two. The rod-shaped particles are characteristically curved in a crescent shape (Torovirus) [13]. The particles usually have large protrusions on the surface in the form of mace or petals (peplomeres or spines). The nucleocapsids have a helical shape and can be detached from the viral particle when treated with detergents. While coronaviruses have a loose nucleocapsid structure, toroviruses have a characteristic tubular shape [14].
In terms of genome size and genetic complexity, coronaviruses are the largest RNA viruses discovered to date. Only okaviruses, large invertebrate nidoviruses from the Roniviridae family, can match them in these characteristics. The replication mechanism has been studied in detail only for the coronaviruses. However, the apparently limited data on toro- and bafiniviruses suggest that they share similar biological characteristics. Virions of these viruses attach to special receptors on the host cell surface using spikes and release their genome into the target cell through fusion of the viral envelope with the plasma membrane and/or the restrictive membrane of the endocytic vesicle [13].
The study of bats as possible vectors of coronaviruses began after the emergence of SARS cases in 2002–2004 [15], and was intensified after the discovery by W. Li et al. detected RNA of Rhinolophus genus (R. pearsoni, R. pussilus, R. macrotis) coronavirus in China, which is genetically close to SARS-CoV [16]. Later, several groups of researchers showed that markers of different coronaviruses (RNA, specific antibodies) in bats are found in North and South America, Europe, and Africa [17–20]. After the outbreak of Middle East respiratory syndrome in 2012–2015, caused by the MERS-CoV coronavirus and affecting several countries, RNA from genetically related coronaviruses was found in bats in Africa and Europe [21, 22]. A SARS-CoV-2-like virus related to COVID-19 was identified in R. affinis in China several years before the pandemic of a new coronavirus infection [23]. The discovery of genetic material of coronaviruses related to human pathogens of severe diseases (SARS-CoV, MERS-CoV, SARS-CoV-2) in bats gave grounds to consider certain species of these animals as potential hosts of precursors of highly pathogenic human coronaviruses.
Physical and geographical characteristics of the analyzed regions
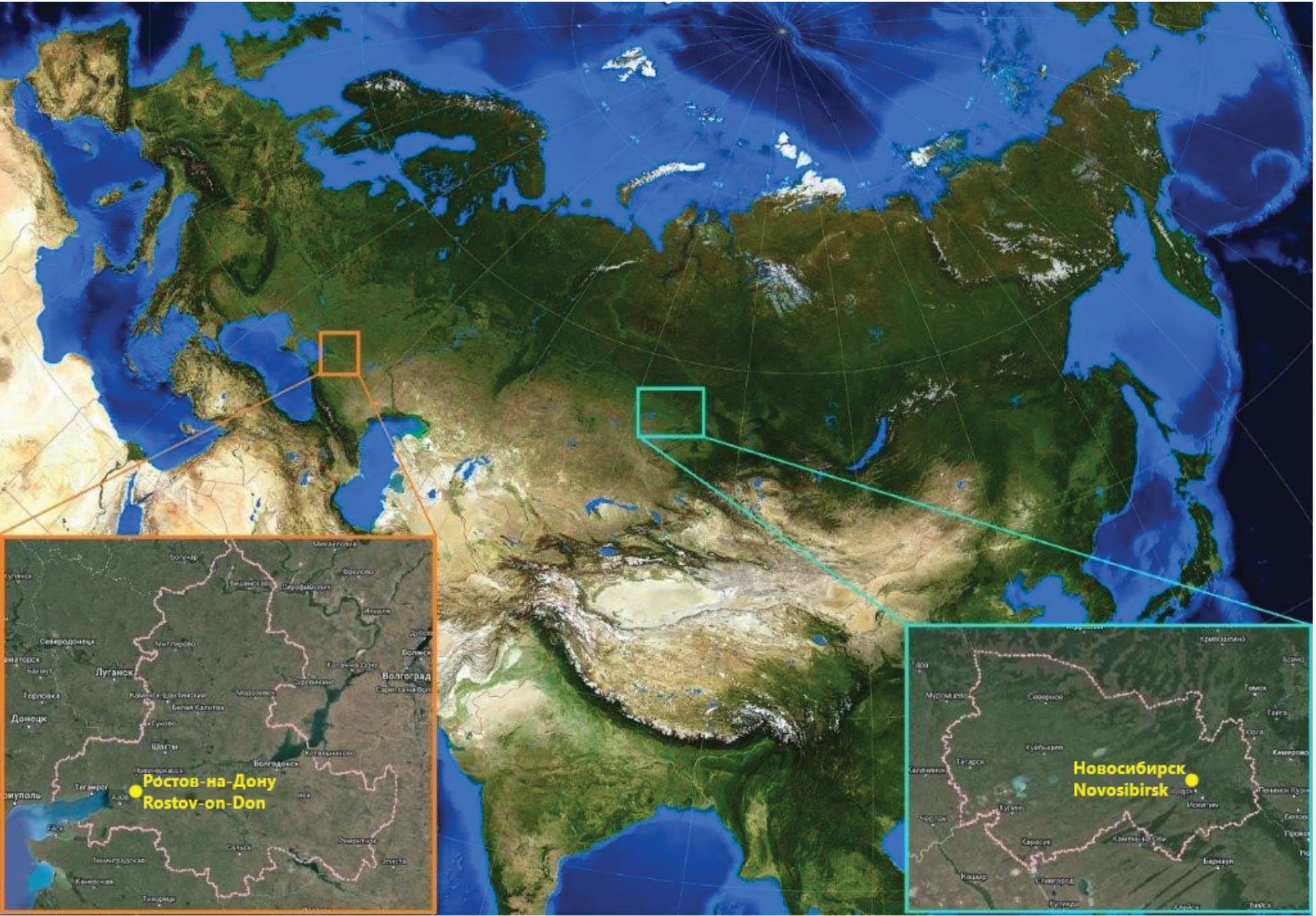
South of European Russia. The administrative center of the Southern Federal District is Rostov region (hereinafter – RR), which is located between 0°14’ – 45°51’ N and 38°14’ – 44º20’E (Figure), in the Lower Don basin, includes the southwestern part of the Russian Plain and the northwestern part of the Precaucasus. In the extreme south-west it is washed by the waters of the Taganrog Bay of the Sea of Azov, in the east by the Tsimlyanskoye Reservoir. Three zonal types of natural landscapes are spread on the territory of the RR: steppe, dry-steppe and semi-desert, which replace each other from west to east. The steppe type of landscapes occupies almost the whole territory of the region, except for its south-eastern part. The dry-steppe type of landscapes occupies the eastern part of the Dono-Chir interfluve and the middle part of the Salo-Manych plain, extending westward along the valley of the Manych River. The semi-desert type of landscapes is confined to the slopes and floodplain terraces of the rivers of the eastern edge of the Salo-Manych Plain [12].

Figure. Geographical location of the analyzed regions on the map of Eurasia, and their administrative centers.
Orange frame – Rostov region, light blue frame – Novosibirsk region.
Рисунок. Географическое положение анализируемых регионов на карте Евразии, их административные центры.
Оранжевая рамка – Ростовская область, светло-голубая рамка – Новосибирская область.
The relief is mainly flat. In the west, there are the eastern spurs of the Donetsk ridge; in the north, there is the Don ridge; in the southeast, there are the slopes of the Ergeni upland; in the south there is the Kumo-Manych depression. The surface of the region is crossed by river valleys, ravines and gullies. The main river is the Don with its tributaries. Forest cover is 2.5% [13]. Artificial (anthropogenic) forms of relief are also widespread, especially on irrigated lands. These are permanent and temporary irrigation canals, spoil heaps, mounds. According to the Köppen-Geiger Climate Classification Map portal (https://www.koppen-map.com/), the climate of most parts of the RR is classified as Dfa (cold continental, without dry season, hot summer), in the northern parts of the region the climate is classified as the Dfb type (cold continental, without dry season, warm summer).
Western Siberia. The Novosibirsk Region (hereinafter referred to as the NSR), as the most climatically and geographically characteristic territory of the Siberian Federal District, is located between 57°14’ – 53°17’ N and 75°05’ – 85°07’ E (Fig. 1), at the junction of the Altai-Sayan mountainous country and the West Siberian Plain, the border between which runs approximately along the Ob River. The Ob River valley divides its territory into two unequal parts. The left-bank part, which makes up about 80% of the territory, is located on the vast plain of the Ob-Irtysh interfluve (Vasyugan plain, Barabinskaya and Kulundinskaya lowlands). Gently sloping uplands alternate here with vast lowlands, poorly dissected by a sparse network of wide flat-bottomed and shallow valleys with slow-flowing rivers. The relief includes ridges and inter-ridge depressions stretching from northeast to southwest for tens of kilometers. There are swamps and lakes in the depressions. The main river is the Ob. The NSR is located in the zones of southern taiga, mixed forests and forest-steppe, with a forest cover of 26% [24]. In the north of the region, there is the southern part of the Vasyugan Plain – the watershed between the tributaries of the Irtysh and Ob, the southern part of the world’s most extensive system of West Siberian high bogs. The western and central part of the region is occupied by the Barabinsk lowland with its characteristic mane relief, numerous drainless lakes and lowland bogs. The south-western and southern parts of the NSR (Kulunda plain) are characterized by a combination of valleys and closed hollows, lakes with strongly mineralized water are widespread. The right-bank (eastern) part of the NSR is characterized by more dissected and elevated relief. Here begin the spurs of the Salair Ridge – a system of valleys, covered with forest, cut by deep beds of numerous small rivers and streams [25]. The climate of most of the NSR is classified as the Dfb type (cold continental, without dry season, warm summer). In the northern parts of the region (the zone of subtaiga forests) the climate type is classified as Dfc (cold continental, without dry season, cold summer), in the southwest, in the steppes of the Kulunda Plain the climate of some areas is classified as BSk (dry steppe cold) (https://www.koppen-map.com/).
Thus, Western Siberia and the south of European Russia are located within the temperate climate belt of Northern Eurasia, but differ significantly in mean annual temperatures, landscape diversity, and the degree of anthropogenic transformation of natural biocenoses. These differences, in turn, will influence the fauna of bats in these regions.
The fauna of mammals of Western Siberia and the south of European Russia
South of European Russia. As of 2013, 15 species of bats belonging to the plain-nosed bat family have been documented in the south of European Russia [4, 7], which is 33% of the species of plain-nosed bats in the fauna of Russia. Sedentary species include all mouse-eared bats (Myotis daubentonii, M. mystacinus, M. dasycneme, M. aurascens), Kuhl’s pipistrelle (Pipistrellus kuhlii), serotine bat (Eptesicus serotinus), and brown long-eared bat (Plecotus auritus) [4]. In the period from May to September in different years, brood colonies of Nathusius’ pipistrelle (P. nathusii), common pipistrelle (P. pipistrellus), two-flowered common pipistrelle (P. pipistrellus), parti-colored bats (Vespertilio murinus), red-headed vesper (Nyctalus noctula), as well as adults of the soprano pipistrelle (P. pygmaeus), northern pipistrelle (E. nilssonii), lesser (N. leisleri) and giant (N. lasiopterus) vesper during seasonal migrations were recorded in the territory of RR [4].
The landscape distribution of bats in the Southern Federal District is heterogeneous due to the presence (or absence) of suitable places for daytime shelters, breeding and hibernation (tree hollows, natural and artificial caves, dwellings and households) and feeding grounds (temporary and permanent freshwater bodies where insects hatch). The highest frequency of occurrence during the activity period is observed in the steppe zone, where representatives of all 15 species of bats were recorded [7], the most numerous of which are the lesser and pond mouse-eared bat, red-headed vesper, Kul’s, pygmy and lesser pipistrelles. The high abundance and species diversity of bats in this zone is associated with the presence of natural forests (12% forest cover in the zone) and human settlement (presence of residential and household buildings). In the dry-steppe zone there are Kul’s pipistrelle, dwarf bat, parti-colored and serotine bats, red-headed vesper, the number of animals is lower than in the steppe zone. In the semi-desert zone, which is the least favorable for mammals, few sightings of the Kul’s pipistrelle and serotine bat were reported [7]. Out of 15 species of bats, representatives of 12 species were recorded in artificial structures, including human dwellings (except for the Lesser and Giant Vespers and the Steppe Nightjar) [7]. The synanthropic species include the Kul’s pipistrelle and the parti-colored bats [4].
Published data on hibernation of bats in the south of European Russia are sketchy. It is reliably known about hibernation of the Red Vesper [26] and the Kul’s pipistrelle [27], the latter prefers to hibernate exclusively in human buildings and may emerge from hibernation during thaws. S.V. Gazaryan et al. with reference to earlier studies report winter findings of the pond, Daubenton’s (M. daubentonii) and whiskered (M. mystacinus) mouse-eared bat, serotine and brown long-eared bat on the territory of the RR [6].
Western Siberia. The chiropterofauna of Siberia includes 10 species of plain-nosed bats (about 22% of Vespertilionidae species in Russia). Information on the species composition of bats in Western Siberia is mainly based on the IUCN Red List of Threatened Species (www.iucnredlist.org) due to the extremely fragmentary data in the scientific literature. The main data on species composition relate to the wintering sites of bats in the caves of the Salair Ridge. A.D. Botvinkin et al. provide data on the presence of 5 species of mouse-eared bats in four caves of the Salair Ridge, located within the NSR: pond (M. dasycneme), eastern (M. petax), Siberian (M. sibirica), Ikonnikov’s (M. ikonnikovi) and long-tailed (M. longicaudatus); Siberian pipistrelle (M. hilgendorfi) and Siberian long-eared bats (P. ognevi) for the observation period from 1978 to 2009 [7]. The presence of pond, eastern, Siberian mouse-eared bat and Siberian long-eared bat was observed in the caves in different seasons of the year, while the Ikonnikov’s mouse-eared bat, long-tailed and Siberian pipistrelle were observed only in winter. The number of animals in caves can vary depending on different factors – year of observation, season and cave size. The largest wintering colony of bats is located in Barsukovskaya Cave (54°22′14′′N 83°57′42′′E) and can number up to several hundred hibernating animals [7]. There are no data on the species distribution and activity period in the steppe, forest-steppe and forest zones of the NSR in the available literature. According to www.iucnredlist.org, the range of the red-headed vesper in the NSR is located in the southern, south-eastern and eastern regions, the animals are found only in summer. The whole territory of the NSR is within the ranges of the parti-colored and northern bats (www.iucnredlist.org). Regarding the latter, A.D. Botvinkin et al. described the finding of an adult female at the entrance to Barsukovskaya Cave in April 1981 [7]. The pond, eastern and Siberian mouse-eared bats, and the Siberian long-eared bats can be classified as sedentary species. There are no official statistics on the species composition of bats flying into or hibernating in dwellings in summer in the settlements of the Siberian Federal District. It can be assumed that in summer, the parti-colored bat, which is a synanthropic species in different parts of its range [7], also uses residential and household buildings for brood colonies or daytime shelters.
Thus, it can be concluded that the fauna of bats in the south of European Russia is characterized by a greater species diversity compared to the chiropterofauna of Western Siberia. Most of the species are specific to the RR or NSR, with the pond bat, common noctule, particolored bat and northern bat being recorded in both regions. Species diversity of bats in southwestern Russia is due to a more favorable temperature regime for feeding, breeding, and hibernation, despite the presence of large open spaces without potential shelter sites. At the same time, the number of hibernating bat species is similar for both regions, despite the harsher temperature regime of the hibernation period in Siberia. In contrast to the RR, where some species of bats (P. kuhlii and N. noctula) are known to hibernate in populated areas, all species hibernating in the NSR form hibernating colonies in caves of the Salair Ridge, which makes these locations unique “incubators” that support hibernation for hundreds of animals for several months. In general, the species diversity of Chiroptera representatives in the regions under consideration shows a dependence on geographic zones and is consistent with the model of global distribution of bats described earlier [28].
Virus-carrying potential in plain-nosed bats inhabiting Western Siberia and southern European Russia
Currently, representatives of 24 families of RNA-containing and 11 families of DNA-containing vertebrate viruses have been identified or isolated in bats [1]. The diversity and number of virus species associated with the Chiroptera order is heterogeneous and actually coincides with the distribution of host animals – the greatest diversity of bat-borne viruses is characteristic of the equatorial, subequatorial, tropical, and subtropical zones [2, 28]. The following viruses or their markers (RNA, viral antigens, specific antibodies) are relevant for public health and frequently occurring in Eurasian temperate zone bats: lyssaviruses (Rhabdoviridae: Lyssavirus), alpha- and betacoronaviruses (Coronaviridae: Alphacoronavirus, Betacoronavirus), including SARS-CoV- and MERS-CoV-like, as well as astroviruses (Astraviridae: Mamastrovirus), paramyxoviruses (Paramyxoviridae), reoviruses (Spinareoviridae: Orthoreovirus), hepeviruses (Hepeviridae: Chirohepevirus), flaviviruses (Flaviviridae: Flavivirus), nairoviruses (Nairoviridae), hantaviruses (Hantaviridae) and influenza A viruses (Orthomyxoviridae: Alphainfluenzavirus) [2, 10–13, 29]. For the species of the West Siberian and southern European part of the country, we searched for information in open sources about cases of identification of topical RNA-containing viruses or their markers, such as lyssaviruses and coronaviruses, in them during the available observation period, because of their greatest relevance and the need for in-depth study (Table 1). Moreover, all bat species mentioned in the subsection belong to the plain-nosed bat family.
Table. Viruses identified in species of plain-nosed bats, distributed in Siberia and southern European Russia bats in Siberia and southern European Russia [2, 9–13, 29]
Таблица. Вирусы, идентифицированные у видов гладконосых летучих мышей, распространенных в Сибири и на юге европейской части России [2, 9–13, 29]
Bat species Виды летучих мышей | Lyssavirus (Rhabdoviridae) | Coronaviridae | Other viruses Другие вирусы | ||||||||||||||||||
KHULV | EBLV-2 | EBLV-1 | RABV | Lissavirus sp. | IRKLV | Alphacoronavirus | Betacoronavirus | MERS-CoV-related | Mamastrovirus | Orthoreovirus sp. | Chirohepevirus | Paramyxovirinae | Yokose virus | Sokuluk virus | Usutu virus | Issyk-Kul virus | Influenza A H3N2 virus | Brno loanvirus | Ledentevirus-like | ||
South of the European part of Russia Юг европейской части России | M. daubentonii | + | + | + | + | + | + | + | + | ||||||||||||
M. mystacinus | + | + | + | + | |||||||||||||||||
M. aurascens | + | ||||||||||||||||||||
P. kuhlii | + | + | + | + | + | + | |||||||||||||||
P. nathusii | + | + | + | + | + | + | |||||||||||||||
P. pipistrellus | + | + | + | + | + | + | + | + | |||||||||||||
P. pygmaeus | + | + | + | ||||||||||||||||||
N. leisleri | + | + | |||||||||||||||||||
N. lasiopterus | + | + | |||||||||||||||||||
P. auritus | + | + | + | + | |||||||||||||||||
E. serotinus | + | + | + | + | + | + | + | ||||||||||||||
Common Общие виды | V. murinus | + | + | + | + | + | + | ||||||||||||||
N. noctula | + | + | + | + | + | + | + | + | + | + | |||||||||||
M. dasycneme | + | + | |||||||||||||||||||
E. nilssonii | + | + | + | ||||||||||||||||||
Siberia Сибирь | M. petax | + | + | + | + | ||||||||||||||||
M. sibirica | + | ||||||||||||||||||||
M. ikonnikovi | + | ||||||||||||||||||||
M. longicaudatus | |||||||||||||||||||||
M. hilgendorfi | + | + | |||||||||||||||||||
P. ognevi | + | ||||||||||||||||||||
European bat lyssavirus 1 (EBLV-1) is widely distributed in the serotine bat E. serotinus population in continental Europe [13], and in 2018 its viral RNA was detected for the first time in E. serotinus in the UK [30]. EBLV-1 is the most common lyssavirus in Europe after rabies virus; the GenBank database contains several dozen sequences of the full genome of the virus from different countries, including Russia (MF187867, LT839613) [24, 25, 31]. According to the World Health Organization, the first case of lyssavirus infection in bats in Europe was registered in Germany in 1954 [32]. In addition to serotine bats, which are considered the natural host of EBLV-1, markers of viral infection (RNA, antigen, specific antibodies) have been detected at different times in several species of bats [13, 31], with virus isolates available in GenBank known only for E. serotinus [24]. M.A. Selimov et al. obtained isolates of Yuli-like virus (EBLV-1) from the common noctule and parti-colored bat in Ukraine [33], indicating that other species of bats may be involved in the maintenance of EBLV-1 circulation. The virus is pathogenic for bats, humans and some domestic animals. With respect to the natural host E. serotinus, cases of mass mortality in brood colonies [34], asymptomatic virus carriage with seroconversion [35] and sporadic mortality of individuals have been described [36]. Experimental infection of late leatherbacks with genetic variant EBLV-1a by intracerebral and subcutaneous injection of the virus led to the development of the disease manifested as food refusal, tremor, irritability, aggressiveness, paralysis, leading to the death of the animal [37]. L. Rønsholt et al. described a case of EBLV-1 isolation from Egyptian fruit bats (Rousettus aegyptiacus) that died with signs of rabies infection 9 days after moving from Rotterdam Zoo (Holland) to Denmark [38]. W.H. Van der Poel et al. showed the pathogenicity for Egyptian fruit bats of EBLV-1 isolates from both R. aegyptiacus and natural hosts of E. serotinus at different routes of infection [39]. I.V. Kuzmin et al. showed the pathogenicity of the Russian strain Yuli (EBLV-1) for common pipistrelles when experimentally infected both when active and when emerging from the winter hibernation in which they were immersed after infection [40]. North American big brown serotine bats E. fuscus demonstrated susceptibility to two strains of EBLV-1a by different routes of infection [41]. The virus is pathogenic to humans; three human infections have been documented during the entire surveillance period – two in 1977 (Ukraine) [42] and 1985 (Russia) [33, 43], and one in France in 2019 [44]. Infection caused by EBLV-1 in humans manifested as progressive encephalitis with signs of hydrophobia [33], paralysis, salivary stasis, and dysphonia [44], and resulted in the death of the diseased person. In 2 cases, a link with bats has been established: in a case from Russia, an 11-year-old girl was infected after being bitten in the lip by a bat [33], In 2 cases, a link with bats has been established: in a case from Russia, an 11-year-old girl was infected after being bitten in the lip by a bat [44]. Cases of EBLV-1 infection have also been noted in the following domestic animals – domestic sheep in Denmark [45], stone marten in Germany [46] and domestic cats in France [47].
European bat lyssavirus 2 (EBLV-2) was first isolated in Finland in 1985 from the brain of a 30-year-old man who died of encephalitis with signs of rabies infection. The patient was a biologist who worked with bats and had a history of multiple bites from these animals [48]. Subsequently, the virus or its markers were detected in pond and Daubenton’s bats in several countries in Western and Northern Europe [49]. There is also a report on the identification of EBLV-2 in M. daubentonii in Ukraine in 1996 [50], which is probably the easternmost point of detection of the virus. In contrast to EBLV-1, isolation of EBLV-2 or detection of its markers during the entire observation period was characteristic only for these two mouse-eared bat species [13, 49], indicating species specificity of the virus. All described cases of EBLV-2 in the natural hosts M. dasycneme and M. daubentonii refer to single individuals found dead or sick [51]. Experimental infection with EBLV-2 in Daubenton’s bats resulted in the development of disease and death of animals only by intracerebral infection; by other routes of infection the virus demonstrated low pathogenicity for natural hosts [52]. During the entire observation period since 1985, 2 cases of fatal encephalitis in humans caused by EBLV-2 have been described. In both cases, the disease was preceded by contact with bats [48, 49]. No cases of EBLV-2 infection of wild and domestic animals in endemic areas have been reported.
Intensive study of lyssaviruses in Central Eurasian mammals began in the 1980s [43]. The first report refers to the isolation of rabies virus from a particolored bat in Western Siberia [53], in which pathogenicity was investigated for common pipistrelles and mongrel white mice [40]. Later, V.A. Ternovoĭ et al. confirmed the presence of rabies virus RNA in brain samples of the eastern mouse-eared bat M. petax (DQ860254, DQ860249, DQ860255) and Ognev’s long-eared bat P. ognevi (DQ860256) collected at wintering sites in the NSR in 2001-2002 [54]. A.I. Karagulov et al. detected rabies virus RNA in brain samples of V. murinus and E. serotinus from 6 regions of Kazakhstan, including the North Kazakhstan region, geographically and ecologically close to the southern regions of Western Siberia [55]. For one sample from E. serotinus it was possible to determine the nucleotide sequence of the N region of the gene (OP585396) of the rabies virus, which is a confirmation of its circulation among bats of this species. Thus, isolation of the virus and repeated detection of its RNA in representatives of several species of Central Eurasian bats allow us to expand our understanding of the role of these animals in the circulation of rabies virus in this region.
The Khujand (KHULV) and Irkut (IRKLV) lyssaviruses are associated with the whiskered mouse-eared bat and Siberian pipistrelle species considered in this work, respectively [43]. Khujand lyssavirus was isolated in 2001 in the vicinity of Khujand (Tajikistan) from a female whiskered mouse-eared bat with signs of lethargy [56]. This is the only report on the isolation of the virus. No cases of disease caused by KHULV in humans or animals were noted in subsequent years. Lyssavirus Irkut was isolated from a male Siberian vesper in 2002 in Irkutsk (Russia). The animal flew into an apartment and was kept for several days, during which it developed weakness, refusal of food and water, leading to subsequent death [57]. Y. Liu et al. IRKLV was isolated from the brain of a clinically healthy male large vesper M. leucogaster captured in 2012 in Jilin Province, China [42]. The virus is pathogenic for humans – G.N. Leonova et al. described a case of fatal lyssavirus encephalitis in a 20-year-old girl in 2007 in Primorsky Krai (Russia). The patient had a history of lip injury from a bat about a month before the onset of the disease. Ozernoe virus was isolated from the brain of the deceased, the sequence of its complete genome (FJ905105) showed 92% homology with the genome of IRKLV isolated from M. hilgendorfi [58]. In 2017. IRKLV RNA was detected in the brain of a deceased dog in Fuxin, Liaoning Province, China [58]. Experimental intramuscular infection with the Chinese IRKV-THChina12 strain of domestic dogs and cats showed low (20% lethality) pathogenicity of IRKLV for dogs and medium (60% lethality) for cats [59]. A serologic survey of bats from Central Europe (Czech Republic, Poland) and Russia (Altai Krai) conducted by V. Seidlova et al. revealed the presence of antibodies to lyssaviruses in representatives of species in which lyssaviruses had been previously isolated or viral RNA had been detected [60]. Seropositive individuals were found among N. noctula in the Czech Republic, among Russian V. murinus, M. hilgendorfi, and M. petax, and for the first time among Siberian mouse-eared bats (M. sibirica). The M. petax and M. sibirica bats belong to sedentary species, therefore, the detection of lyssavirus infection markers in them indicates the circulation of lyssaviruses in their West Siberian population.
Numerous alpha- and betacoronaviruses have now been identified in bats in different regions of the world. The plain-nosed Vespertilionidae have the highest frequency of alphacoronavirus RNA in Europe and Asia, while for betacoronaviruses they are the second most frequently detected viral RNA after the horseshoe bats of Rhinolophidae in Europe, and the third after Rhinolophidae and the wings of Pteropodidae in Asia [61]. Within the genus Alphacoronavirus, viruses from 9 of 14 subgenera have been identified in bats, with the subgenera Colacovirus, Pedacovirus, Nyctacovirus and Myotacovirus associated with smooth-footed boobies. Within Betacoronavirus, 4 out of 5 subgenera were associated with bats, of which only Merbecovirus was found in plain-nosed bats [62].
The carriage of coronaviruses among European species of bats is well studied. C. Kohl et al. have cited data on detection of alpha- and betacoronavirus RNA in bats in different European countries over several years of observation [10]. Genetic material of alpha- and betacoronaviruses, including SARS-CoV- and MERS-CoV-like viruses, has also been detected in bats from the fauna of Russia [63‒67]. RNA of alpha-coronaviruses was detected in fecal samples of P. kuhlii captured in Rostov-on-Don in 2021 [64]. Coronavirus RNA (without identification to genus) was also detected in oropharyngeal swabs of P. kuhlii and E. serotinus in the neighboring regions of the Rostov region – the Republic of Adygea and Krasnodar Krai [65], which indicates the circulation of coronaviruses among bats in southern Russia. According to literature data, MERS-CoV-like viruses have been associated with nuthatches in several European countries, including Russia [21, 22, 66]. The bats of central Eurasia are insufficiently studied in this respect. A.I. Karagulov et al. detected alphacoronavirus RNA in fecal samples of V. murinus and E. serotinus in the western and southern regions of Kazakhstan [55].
Relatively recently, alphacoronavirus RNA was identified in eastern and Ikonnikov’s mouse-eared bats (M. ikonnikovi) in southern Western Siberia [67], indicating the need for further studies of coronaviruses in populations of bats in this region.
All the above mentioned cases of coronavirus detection in bats in Europe and central Eurasia concern only detection of viral RNA without isolation of live viruses. The pathogenicity of the identified coronaviruses to their natural hosts has not been established. A. Moreno et al. identified the complete genome of a MERS-CoV-like virus in fresh cadavers of P. kuhlii and H. savii obtained from the Bat Rehabilitation Center [22], but it cannot be stated that MERS-like coronavirus was the cause of death in these cases. Among terrestrial animals in Eurasia, only in hedgehogs (Erinaceus europaeus, E. amurensis), genetically similar MERS-like coronaviruses have been identified during the entire observation period [68–70] without any clinical manifestations. No cases of human infection with bat-borne coronaviruses have been described in Europe or central Eurasia. However, given the diversity of identified coronaviruses, including MERS-CoV-like ones in migratory (P. nathusii) and synanthropic (P. kuhlii) species of bats, it is important to monitor these viruses in populations of bats both in Western Siberia and the south of European Russia.
Conclusion
Thus, circulation of representatives of such viral genera and families as lyssaviruses, alpha- and betacoronaviruses, astroviruses, paramyxoviruses, reoviruses, hepeviruses, flaviviruses, nairoviruses, hantaviruses and influenza A viruses has been detected in plain-nosed bats in European Russia, of which lethal cases in the human population were established for European bat lyssavirus EBLV-1. The presence of EBLV-1 and IRKLV lyssaviruses was established in plain-nosed bats in the territories of Western Siberia. Moreover, IRKLV was isolated from the big pipistrelle Murina leucogaster (Irkutsk). Alphacoronaviruses were detected among bats both in Western Siberia and European Russia. However, in addition to alphacoronaviruses, betacoronaviruses, including SARS-CoV- and MERS-CoV-like viruses, have also been identified in European territories. In general, the number of detected viruses in plain-nosed bats in the south of European Russia is higher than in Siberia, which suggests a higher potential for virus carriage. The proven presence of such viruses dangerous for humans as European bat lyssavirus in the populations of the plain-nosed bat family in these regions indicates the existence of the danger of natural foci of infection. In this regard, further systematic studies of the species composition and distribution of genetic variants of lyssaviruses and coronaviruses in plain-nosed bats in these areas are necessary.
As of 2024, mammals of the Vespertilionidae family are natural reservoirs or vectors of a number of viruses that are dangerous for humans. At the same time, in the Russian Federation, the epidemic potential of viruses detected in these animals has been studied insufficiently and fragmentarily, which does not allow for reliable forecasting of the situation. The approach to monitoring and assessment of existing epidemiological threats should be revised, especially in connection with the pandemic of a new coronavirus infection (COVID-19), which was caused by a fact that was well-known, but insufficiently studied earlier.
About the authors
Olesia V. Ohlopkova
Central Research Institute of Epidemiology, Rospotrebnadzor; Virology Research Institute of the Federal Research Center for Fundamental and Translational Medicine
Author for correspondence.
Email: ohlopkova.lesia@yandex.ru
ORCID iD: 0000-0002-8214-7828
PhD in Biology, Senior Researcher of the Laboratory of Experimental Pharmacology
Россия, 111123, Moscow; 630060, NovosibirskYulia V. Kononova
Virology Research Institute of the Federal Research Center for Fundamental and Translational Medicine
Email: yuliakononova07@yandex.ru
ORCID iD: 0000-0002-3677-3668
PhD in Biology, Senior Researcher
Россия, 630060, NovosibirskMarina A. Tyumentseva
Central Research Institute of Epidemiology, Rospotrebnadzor
Email: tyumantseva@cmd.ru
ORCID iD: 0000-0002-3145-3702
PhD in Biology, Head of the Laboratory of Genomic Editing of the Department of Molecular Diagnostics and Epidemiology
Россия, 111123, MoscowAlexander I. Tyumentsev
Central Research Institute of Epidemiology, Rospotrebnadzor
Email: tyumantsev@cmd.ru
ORCID iD: 0000-0003-0537-2586
PhD in Biology, Head of the Laboratory of Experimental Pharmacology, Department of Molecular Diagnostics and Epidemiology
Россия, 111123, MoscowAlexander M. Shestopalov
Virology Research Institute of the Federal Research Center for Fundamental and Translational Medicine
Email: amshestopalov@frcftm.ru
ORCID iD: 0000-0002-9734-0620
Doctor of Biological Sciences, Professor, Honored Worker of Science of the Russian Federation, Director
Россия, 630060, NovosibirskVasily G. Akimkin
Central Research Institute of Epidemiology, Rospotrebnadzor
Email: akimkin@pcr.ms
ORCID iD: 0000-0003-4228-9044
Doctor of Medical Sciences, Academician of the Russian Academy of Sciences, Professor, Honored Doctor of the Russian Federation, Laureate of the Prize of the Government of the Russian Federation in the field of science and technology, Director
Россия, 111123, MoscowReferences
- Van Brussel K., Holmes E.C. Zoonotic disease and virome diversity in bats. Curr. Opin. Virol. 2022; 52: 192–202. https://doi.org/10.1016/j.coviro.2021.12.008
- Liu Z., Liu Q., Wang H., Yao X. Severe zoonotic viruses carried by different species of bats and their regional distribution. Clin. Microbiol. Infect. 2024; 30(2): 206–10. https://doi.org/10.1016/j.cmi.2023.09.025
- Poleshchuk E.M., Tagakova D.N., Sidorov G.N., Orlova T.S., Gordeiko N.S., Kaisarov A.Z. Lethal cases of lyssavirus encephalitis in humans after contact with bats in the Russian Far East in 2019–2021. Voprosy virusologii. 2023; 68(1): 45–58. https://doi.org/10.36233/0507-4088-156 https://elibrary.ru/bjmlbx
- Sidorchuk A.A. Book Review: Makarov V.V., Gulyukin A.M., Gulyukin M.I. Rabies: natural history at the turn of the century. Moscow: ZooVetKniga; 2015. Voprosy virusologii. 2018; 63(1): 48. https://elibrary.ru/yoomce (in Russian)
- Alekseeva S.V. Book Review: Makarova V.V., Lozovogo D.A. New especially dangerous infections associated with bats. Vladimir; 2016. Veterinariya segodnya. 2017; (2): 67–8. https://elibrary.ru/yrtnpb (in Russian)
- Gazaryan S.V., Bakhtadze G.B., Malinovkin A.V. Current state of knowledge of bats in Rostov region. Plecotus et al. 2010; (13): 50–8. https://elibrary.ru/wcnffh (in Russian)
- Botvinkin A.D., VakhrushevA.V., Kuz’min I.V., Morozov P.N., Shevyrnogov S.Z. From the history of bat observations in caves of Salair mountain ridge (Novosibirsk region). Plecotus et al. 2015; (18): 45–53. https://elibrary.ru/vucerz (in Russian)
- Arai S., Yanagihara R. Genetic diversity and geographic distribution of bat-borne hantaviruses. Curr. Issues. Mol. Biol. 2020; 39: 1–28. https://doi.org/10.21775/cimb.039.001
- Wang B., Yang X.L. Chirohepevirus from bats: insights into hepatitis E virus diversity and evolution. Viruses. 2022; 14(5): 905. https://doi.org/10.3390/v14050905
- Kohl C., Nitsche A., Kurth A. Update on potentially zoonotic viruses of European bats. Vaccines (Basel). 2021; 9(7): 690. https://doi.org/10.3390/vaccines9070690
- Johnson N., Cunningham A.F., Fooks A.R. The immune response to rabies virus infection and vaccination. Vaccine. 2010; 28(23): 3896–901. https://doi.org/10.1016/j.vaccine.2010.03.039
- Wambugu E.N., Kimita G., Kituyi S.N., Washington M.A., Masakhwe C., Mutunga L.M., et al. Geographic distribution of rabies virus and genomic sequence alignment of wild and vaccine strains, Kenya. Emerg. Infect. Dis. 2024; 30(8): 1642–50. https://doi.org/10.3201/eid3008.230876.
- Maier H.J., Bickerton E., Britton P. Preface. Coronaviruses. Methods Mol. Biol. 2015; 1282: v. https://doi.org/10.1007/978-1-4939-2438-7
- de Vries A.A.F., Horzinek M.C., Rottier P.J.M., de Groot R.J. The genome organization of the nidovirales: similarities and differences between Arteri-, Toro-, and Coronaviruses. Semin. Virol. 1997; 8(1): 33–47. https://doi.org/10.1006/smvy.1997.0104
- Poon L.L., Chu D.K., Chan K.H., Wong O.K., Ellis T.M., Leung Y.H., et al. Identification of a novel coronavirus in bats. J. Virol. 2005; 79(4): 2001–9. https://doi.org/10.1128/JVI.79.4.2001-2009.2005
- Li W., Shi Z., Yu M., Ren W., Smith C., Epstein J.H., et al. Bats are natural reservoirs of SARS-like coronaviruses. Science. 2005; 310(5748): 676–9. https://doi.org/10.1126/science.1118391
- Dominguez S.R., O’Shea T.J., Oko L.M., Holmes K.V. Detection of group 1 coronaviruses in bats in North America. Emerg. Infect. Dis. 2007; 13(9): 1295–300. https://doi.org/10.3201/eid1309.070491
- Müller M.A., Paweska J.T., Leman P.A., Drosten C., Grywna K., Kemp A., et al. Coronavirus antibodies in African bat species. Emerg. Infect. Dis. 2007; 13(9): 1367–70. https://doi.org/10.3201/eid1309.070342
- Gloza-Rausch F., Ipsen A., Seebens A., Göttsche M., Panning M., Drexler J.F., et al. Detection and prevalence patterns of group I coronaviruses in bats, northern Germany. Emerg. Infect. Dis. 2008; 14(4): 626–31. https://doi.org/10.3201/eid1404.071439
- Carrington C.V., Foster J.E., Zhu H.C., Zhang J.X., Smith G.J., Thompson N., et al. Detection and phylogenetic analysis of group 1 coronaviruses in South American bats. Emerg. Infect. Dis. 2008; 14(12): 1890–3. https://doi.org/10.3201/eid1412.080642
- Annan A., Baldwin H.J., Corman V.M., Klose S.M., Owusu M., Nkrumah E.E., et al. Human betacoronavirus 2c EMC/2012-related viruses in bats, Ghana and Europe. Emerg. Infect. Dis. 2013; 19(3): 456–9. https://doi.org/10.3201/eid1903.121503
- Moreno A., Lelli D., de Sabato L., Zaccaria G., Boni A., Sozzi E., et al. Detection and full genome characterization of two beta CoV viruses related to Middle East respiratory syndrome from bats in Italy. Virol. J. 2017; 14(1): 239. https://doi.org/10.1186/s12985-017-0907-1
- Zhou P., Yang X.L., Wang X.G., Hu B., Zhang L., Zhang W., et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020; 579(7798): 270–3. https://doi.org/10.1038/s41586-020-2012-7
- Forró B., Marton S., Fehér E., Domán M., Kemenesi G., Cadar D., et al. Phylogeny of Hungarian EBLV-1 strains using whole-genome sequence data. Transbound. Emerg. Dis. 2021; 68(3): 1323–31. https://doi.org/10.1111/tbed.13789
- Selimov M.A., Tatarov A.G., Botvinkin A.D., Klueva E.V., Kulikova L.G., Khismatullina N.A. Rabies-related Yuli virus; identification with a panel of monoclonal antibodies. Acta Virol. 1989; 33(6): 542–6.
- Smirnov D.G., Zabashta A.V. Distribution of the noctule bat (nyctalus noctula) in the Rostov region and the problem of expanding the northern boundary of its wintering area. Rossiiskii zhurnal biologicheskikh invazii. 2023; 14(4): 621–33. https://doi.org/10.35885/1996-1499-16-3-168-183 https://elibrary.ru/raavaq (in Russian)
- Minoranskii V.A.E., Malinovkin A.V. Kul bat (pipistrellus kuhlii kuhl.) in the Rostov region. Izvestiya vysshikh uchebnykh zavedenii. Severo-Kavkazskii region. Seriya: estestvennye nauki. 2015; (2): 80–3. https://elibrary.ru/txjfqr (in Russian)
- Alves D.M.C.C., Diniz-Filho J.A.F., Souza K.D, Gouveia S.F., Villalobos F. Geographic variation in the relationship between large-scale environmental determinants and bat species richness. Basic Appl. Ecol. 2018; 27: 1–8. https://doi.org/10.1016/j.baae.2017.12.002
- Yang W., Schountz T., Ma W. Bat influenza viruses: current status and perspective. Viruses. 2021; 13(4): 547. https://doi.org/10.3390/v13040547
- Folly A.J., Marston D.A., Golding M., Shukla S., Wilkie R., Lean F.Z.X., et al. Incursion of European Bat Lyssavirus 1 (EBLV-1) in serotine bats in the United Kingdom. Viruses. 2021; 13(10): 1979. https://doi.org/10.3390/v13101979
- Harris S.L., Brookes S.M., Jones G., Hutson A.M., Racey P.A., Aegerter J., et al. European bat lyssaviruses: Distribution, prevalence and implications for conservation. Biol. Conserv. 2006; 131(2): 193–210. https://doi.org/10.1016/j.biocon.2006.04.006
- Pérez-Jordá J.L., Ibánez C., Muñoz-Cervera M., Téllez A. Lyssavirus in Eptesicus serotinus (Chiroptera: Vespertilionidae). J. Wildl. Dis. 1995; 31(3): 372–7. https://doi.org/10.7589/0090-3558-31.3.372
- Selimov M.A., Smekhov A.M., Antonova L.A., Shablovskaya E.A., King A.A., Kulikova L.G. New strains of rabies-related viruses isolated from bats in the Ukraine. Acta Virol. 1991; 35(3): 226–31.
- Picard-Meyer E., Servat A., Wasniewski M., Gaillard M., Borel C., Cliquet F. Bat rabies surveillance in France: first report of unusual mortality among serotine bats. BMC Vet. Res. 2017; 13(1): 387. https://doi.org/10.1186/s12917-017-1303-1
- Robardet E., Borel C., Moinet M., Jouan D., Wasniewski M., Barrat J., et al. Longitudinal survey of two serotine bat (Eptesicus serotinus) maternity colonies exposed to EBLV-1 (European Bat Lyssavirus type 1): Assessment of survival and serological status variations using capture-recapture models. PLoS Negl. Trop. Dis. 2017; 11(11): e0006048. https://doi.org/10.1371/journal.pntd.0006048
- Mingo-Casas P., Sandonís V., Obón E., Berciano J.M., Vázquez-Morón S., Juste J., et al. First cases of European bat lyssavirus type 1 in Iberian serotine bats: Implications for the molecular epidemiology of bat rabies in Europe. PLoS Negl. Trop. Dis. 2018; 12(4): e0006290. https://doi.org/10.1371/journal.pntd.0006290
- Freuling C., Vos A., Johnson N., Kaipf I., Denzinger A., Neubert L., et al. Experimental infection of serotine bats (Eptesicus serotinus) with European bat lyssavirus type 1a. J. Gen. Virol. 2009; 90(Pt 10): 2493–502. https://doi.org/10.1099/vir.0.011510-0
- Rønsholt L., Sørensen K.J., Bruschke C.J., Wellenberg G.J., van Oirschot J.T., Johnstone P., et al. Clinically silent rabies infection in (zoo) bats. Vet. Rec. 1998; 142(19): 519–20. https://doi.org/10.1136/vr.142.19.519
- Van der Poel W.H., van der Heide R., van Amerongen G., van Keulen L.J., Wellenberg G.J., Bourhy H., et al. Characterisation of a recently isolated lyssavirus in frugivorous zoo bats. Arch. Virol. 2000; 145(9): 1919–31. https://doi.org/10.1007/s007050070066
- Kuz’min I.V., Botvinkin A.D., Shaĭmardanov R.T. Experimental lyssavirus infection in chiropters. Voprosy virusologii. 1994; 39(1): 17–21.
- Franka R., Johnson N., Müller T., Vos A., Neubert L., Freuling C., et al. Susceptibility of North American big brown bats (Eptesicus fuscus) to infection with European bat lyssavirus type 1. J. Gen. Virol. 2008; 89(Pt 8): 1998–2010. https://doi.org/10.1099/vir.0.83688-0
- Liu Y., Zhang S., Zhao J., Zhang F., Hu R. Isolation of Irkut virus from a Murina leucogaster bat in China. PLoS Negl. Trop. Dis. 2013; 7(3): e2097. https://doi.org/10.1371/journal.pntd.0002097
- Kuzmin I.V., Botvinkin A.D., Poleschuk E.M., Orciari L.A., Rupprecht C.E. Bat rabies surveillance in the former Soviet Union. Dev. Biol. (Basel). 2006: 125: 273–82.
- Regnault B., Evrard B., Plu I., Dacheux L., Troadec E., Cozette P., et al. First case of lethal encephalitis in Western Europe due to European bat lyssavirus type 1. Clin. Infect. Dis. 2022; 74(3): 461–6. https://doi.org/10.1093/cid/ciab443
- Tjørnehøj K., Fooks A.R., Agerholm J.S., Rønsholt L. Natural and experimental infection of sheep with European bat lyssavirus type-1 of Danish bat origin. J. Comp. Pathol. 2006; 134(2-3): 190–201. https://doi.org/10.1016/j.jcpa.2005.10.005
- Müller T., Cox J., Peter W., Schäfer R., Johnson N., McElhinney L.M., et al. Spill-over of European bat lyssavirus type 1 into a stone marten (Martes foina) in Germany. J. Vet. Med. B. Infect. Dis. Vet. Public Health. 2004; 51(2): 49–54. https://doi.org/10.1111/j.1439-0450.2003.00725.x
- Dacheux L., Larrous F., Mailles A., Boisseleau D., Delmas O., Biron C., et al. European bat Lyssavirus transmission among cats, Europe. Emerg. Infect. Dis. 2009; 15(2): 280–4. https://doi.org/10.3201/eid1502.080637
- Lumio J., Hillbom M., Roine R., Ketonen L., Haltia M., Valle M., et al. Human rabies of bat origin in Europe. Lancet. 1986; 1(8477): 378. https://doi.org/10.1016/s0140-6736(86)92336-6
- McElhinney L.M., Marston D.A., Wise E.L., Freuling C.M., Bourhy H., Zanoni R., et al. Molecular epidemiology and evolution of European bat lyssavirus 2. Int. J. Mol. Sci. 2018; 19(1): 156. https://doi.org/10.3390/ijms19010156
- Harris S.L., Brookes S.M., Jones G., Hutson A.M., Racey P.A., Aegerter J., et al. European bat lyssaviruses: Distribution, prevalence and implications for conservation. Biol. Conserv. 2006; 131(2): 193–210. https://doi.org/10.1016/j.biocon.2006.04.006
- Johnson N., Wakeley P.R., Brookes S.M., Fooks A.R. European bat lyssavirus type 2 RNA in Myotis daubentonii. Emerg. Infect. Dis. 2006; 12(7): 1142–4. https://doi.org/10.3201/eid1207.060287
- Johnson N., Vos A., Neubert L., Freuling C., Mansfield K.L., Kaipf I., et al. Experimental study of European bat lyssavirus type-2 infection in Daubenton’s bats (Myotis daubentonii). J. Gen. Virol. 2008; 89(Pt. 11): 2662–72. https://doi.org/10.1099/vir.0.2008/003889-0
- Botvinkin A.D. Rabies virus was isolates from a bat in Western Siberia. Zhurnal mikrobiologii, epidemiologii i immunobiologii. 1988; 65(7): 97–8. https://elibrary.ru/tfsull (in Russian)
- Ternovoi V.A., Zaikovskaia A.V., Tomilenko A.A., Rassadkin Yu.N., Aksenov V.I., Chausov E.V., et al. Lissavirus in bats residing in the South of West Siberia. Voprosy virusologii. 2005; 50(1): 31–4. https://elibrary.ru/hrxxvd (in Russian)
- Karagulov A.I., Argimbayeva T.U., Omarova Z.D., Tulendibayev A.B., Dushayeva L.Z., Svotina M.A., et al. The prevalence of viral pathogens among bats in Kazakhstan. Viruses. 2022; 14(12): 2743. https://doi.org/10.3390/v14122743
- Kuzmin I.V., Orciari L.A., Arai Y.T., Smith J.S., Hanlon C.A., Kameoka Y., et al. Bat lyssaviruses (Aravan and Khujand) from Central Asia: phylogenetic relationships according to N, P and G gene sequences. Virus Res. 2003; 97(2): 65–79. https://doi.org/10.1016/s0168-1702(03)00217-x
- Botvinkin A.D., Poleschuk E.M., Kuzmin I.V., Borisova T.I., Gazaryan S.V., Yager P., et al. Novel lyssaviruses isolated from bats in Russia. Emerg. Infect. Dis. 2003; 9(12): 1623–5. https://doi.org/10.3201/eid0912.030374
- Leonova G.N., Somova L.M., Belikov S.I., Kondratov I.G., Plekhova N.G., Krylova N.V., et al. The fatal case of lyssavirus encephalitis in the Russian Far East. Encephalitis. 2013. https://doi.org/10.5772/52869
- Chen T., Miao F.M., Liu Y., Zhang S.F., Zhang F., Li N., et al. Possible transmission of Irkut virus from dogs to humans. Biomed. Environ. Sci. 2018; 31(2): 146–8. https://doi.org/10.3967/bes2018.017
- Seidlova V., Zukal J., Brichta J., Anisimov N., Apoznański G., Bandouchova H., et al. Active surveillance for antibodies confirms circulation of lyssaviruses in Palearctic bats. BMC Vet. Res. 2020; 16(1): 482. https://doi.org/10.1186/s12917-020-02702-y
- Wang Z., Huang G., Huang M., Dai Q., Hu Y., Zhou J., et al. Global patterns of phylogenetic diversity and transmission of bat coronavirus. Sci. China Life Sci. 2023; 66(4): 861–74. https://doi.org/10.1007/s11427-022-2221-5
- Ruiz-Aravena M., McKee C., Gamble A., Lunn T., Morris A., Snedden C.E., et al. Ecology, evolution and spillover of coronaviruses from bats. Nat. Rev. Microbiol. 2022; 20(5): 299–314. https://doi.org/10.1038/s41579-021-00652-2
- Alkhovsky S., Lenshin S., Romashin A., Vishnevskaya T., Vyshemirsky O., Bulycheva Y., et al. SARS-like coronaviruses in Horseshoe bats (Rhinolophus spp.) in Russia, 2020. Viruses. 2022; 14(1): 113. https://doi.org/10.3390/v14010113
- Korneenko E.V., Samoilov A.E., Chudinov I.K., Butenko I.O., Sonets I.V., Artyushin I.V., et al. Alphacoronaviruses from bats captured in European Russia in 2015 and 2021 are closely related to those of Northern Europe. Front. Ecol. Evol. 2024; 12: 1324605. https://doi.org/10.3389/fevo.2024.1324605
- Popov I.V., Ohlopkova O.V., Donnik I.M., Zolotukhin P.V., Umanets A., Golovin S.N., et al. Detection of coronaviruses in insectivorous bats of Fore-Caucasus, 2021. Sci. Rep. 2023; 13(1): 2306. https://doi.org/10.1038/s41598-023-29099-6
- Speranskaya A.S., Artiushin I.V., Samoilov A.E., Korneenko E.V., Khabudaev K.V., Ilina E.N., et al. Identification and Genetic Characterization of MERS-Related Coronavirus Isolated from Nathusius’ Pipistrelle (Pipistrellus nathusii) near Zvenigorod (Moscow Region, Russia). Int. J. Environ. Res. Public Health. 2023; 20(4): 3702. https://doi.org/10.3390/ijerph20043702
- Yashina L.N., Zhigalin A.V., Abramov S.A., Luchnikova E.M., Smetannikova N.A., Dupal T.A., et al. Coronaviruses (Coronaviridae) of bats in the northern Caucasus and south of western Siberia. Voprosy virusologii. 2024; 69(3): 255–65. https://doi.org/10.36233//0507-4088-233 https://elibrary.ru/konejo
- Corman V.M., Kallies R., Philipps H., Göpner G., Müller M.A., Eckerle I., et al. Characterization of a novel betacoronavirus related to middle East respiratory syndrome coronavirus in European hedgehogs. J. Virol. 2014; 88(1): 717–24. https://doi.org/10.1128/JVI.01600-13
- Pomorska-Mól M., Ruszkowski J.J., Gogulski M., Domanska-Blicharz K. First detection of Hedgehog coronavirus 1 in Poland. Sci. Rep. 2022; 12(1): 2386. https://doi.org/10.1038/s41598-022-06432-z
- Lau S.K.P., Luk H.K.H., Wong A.C.P., Fan R.Y.Y., Lam C.S.F., Li K.S.M., et al. Identification of a novel Betacoronavirus (Merbecovirus) in Amur hedgehogs from China. Viruses. 2019; 11(11): 980. https://doi.org/10.3390/v11110980
Supplementary files

