



Вспомогательный белок Vpr вируса иммунодефицита человека 1-го типа (Retroviridae: Orthoretrovirinae: Lentivirus: Human immunodeficiency virus-1): особенности генетических вариантов вируса, циркулировавших на территории Московской области в 2019–2020 гг.
- Авторы: Кузнецова А.И.1, Антонова А.А.1, Макеева Е.А.2, Ким К.В.1, Мунчак Я.М.1, Меженская Е.Н.1, Орлова-Морозова Е.А.3, Пронин А.Ю.3, Прилипов А.Г.1, Галзитская О.В.1,4
-
Учреждения:
- ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
- ФГАОУ ВО «Московский политехнический университет»
- ГБУЗ МО «Центр профилактики и борьбы со СПИД»
- ФБУН «Институт теоретической и экспериментальной биофизики» Российской академии наук
- Выпуск: Том 70, № 4 (2025)
- Страницы: 324-339
- Раздел: ОРИГИНАЛЬНЫЕ ИССЛЕДОВАНИЯ
- URL: https://virusjour.crie.ru/jour/article/view/16720
- DOI: https://doi.org/10.36233/0507-4088-296
- EDN: https://elibrary.ru/mfgcsm
- ID: 16720

Цитировать

Аннотация
Введение. Vpr является многофункциональным вспомогательным белком ВИЧ-1, который проявляет свою активность на разных этапах жизненного цикла вируса. Олигомеризация белка является необходимым условием для проникновения Vpr в вирион и последующего его участия на ранних этапах ВИЧ-инфекции. К настоящему времени определены естественные аминокислотные замены в Vpr, ассоциированные с прогрессированием заболевания; рассматривается возможность создания средств терапии на основе Vpr.
Цель исследования – изучение особенностей Vpr у наиболее распространенных генетических вариантов ВИЧ-1, циркулировавших на территории Московской области в 2019–2020 гг.
Материалы и методы. Исследовали образцы ВИЧ-1, полученные от 231 пациента ГБУЗ МО «Центр профилактики и борьбы со СПИД» в период 2019–2020 гг. с разными стадиями заболевания, по схеме: экстракция провирусной ДНК, амплификация гена vpr, секвенирование, анализ полученных данных. Изучали консенсусные последовательности Vpr наиболее распространенных в России генетических вариантов и их пространственные структуры, вариабельность Vpr вариантов ВИЧ-1 суб-субтипа A6 (Vpr-A6) у пациентов с разными стадиями заболевания.
Результаты. Особенности Vpr различных генетических вариантов ВИЧ-1, циркулирующих в России, могут влиять на формирование его олигомерных форм. Не было выявлено ни одного сайта со статистически значимыми различиями в частоте встречаемости аминокислотных замен у пациентов с разными стадиями заболевания.
Заключение. Белки Vpr генетических вариантов ВИЧ-1, циркулирующих в России, могут иметь различия в функциональных свойствах. Варианты Vpr-A6 имели низкую вариабельность у пациентов с разными стадиями заболевания, в связи с чем Vpr-А6 может рассматриваться как объект для создания средств терапии.
Ключевые слова
Полный текст
Введение
Вспомогательный белок Vpr вируса иммунодефицита человека 1-го типа (Retroviridae: Orthoretrovirinae: Lentivirus: Human immunodeficiency virus-1; ВИЧ-1) является высококонсервативным вирусным белком, состоящим из 96 аминокислотных остатков (а.о.), массой около 14 кДа [1]. В своей структуре белок Vpr содержит три α-спирали, которые образованы участками белковой цепи с 17-го по 33-й а.о., с 38-го по 50-й а.о. и с 55-го по 77-й а.о. (рис. 1). α-Спирали левозакручены и ориентированы друг относительно друга таким образом, чтобы обеспечить взаимодействия между следующими аминокислотными остатками: L20, L23, L26, A30, V31 первой α-спирали, W38, L39, L42, I46 второй α-спирали и V57, L60, I61, L64, L68, F72 третьей α-спирали. Вышеуказанная ориентация дополнительно фиксируется взаимодействиями между T19, L20, W54 с одной стороны и между H33, F34, H71, F72 с другой. Структура белка Vpr характеризуется гибкими N- и C-концевыми областями: с 1-го по 13-й а.о. и с 78-го по 96-й а.о. соответственно [2].

Рис. 1. Схематическое изображение первичной структуры белка Vpr.
M – метионин; E – глутаминовая кислота; D – аспарагиновая кислота; T – треонин; L – лейцин; A – аланин; V – валин; H – гистидин; F – фенилаланин; I – изолейцин; Y – тирозин; W – триптофан; R – аргинин; S – серин; Vpr T-Helper/CD4+ Epitope region (major) – область белка Vpr, в которой преимущественно были картированы эпитопы, распознающиеся иммунной системой для последующего развития CD4+-T-клеточного ответа (https://www.hiv.lanl.gov/content/immunology/maps/helper/Vpr.html); Vpr CTL/CD8+ Epitope region (major) – область белка Vpr, в которой преимущественно были картированы эпитопы, распознающиеся иммунной системой для последующего развития CD8+-цитотоксического Т-клеточного ответа (https://www.hiv.lanl.gov/content/immunology/maps/ctl/Vpr.html).
Fig. 1. Schematic representation of Vpr primary structure.
M – methionine; E – glutamic acid; D – aspartic acid; T – threonine; L – leucine; A – alanine; V – valine; H – histidine; F – phenylalanine; I – isoleucine; Y – tyrosine; W – tryptophan; R – arginine; S – serine; Vpr T-Helper/CD4+ Epitope region (major) – Vpr region in which predominantly the epitopes have been mapped that are recognized by the immune system for subsequent development of CD4+ T cell response; Vpr CTL/CD8+ Epitope region (major) – Vpr region in which predominantly the epitopes have been mapped that are recognized by the immune system for subsequent development of CD8+ cytotoxic T cell response (https://www.hiv.lanl.gov/content/immunology/maps/ctl/Vpr.html).
Ген белка Vpr экспрессируется на поздних стадиях жизненного цикла ВИЧ-1 и связывается с вирусным белком-предшественником Pr55Gag, который играет важную роль в сборке и продукции вирусных частиц. Олигомеризация Vpr имеет решающее значение для распознавания Pr55Gag, затем Pr55Gag-Vpr-комплексы накапливаются в плазматической мембране для последующего эффективного включения в вирионы [3–5]. Таким образом, при инфицировании вирусом клетки-хозяина белок Vpr проникает в нее в составе вириона, что позволяет ему активно участвовать уже в ранних стадиях репликации вируса. Белок Vpr обладает многочисленными функциями (рис. 2):
- повышает процессивность и точность обратной транскрипции [6, 7];
- входит в состав прединтеграционного комплекса (вирусная ДНК, интеграза, белок Vpr и т.д.), который обеспечивает доставку вирусной ДНК из цитоплазмы в ядро для последующей интеграции в геном клетки-хозяина [6, 7];
- усиливает транскрипцию провирусной ДНК [6–8];
- вызывает убиквитин/протеасомозависимую деградацию некоторых клеточных белков, останавливая клеточный цикл в фазе G2, что способствует созданию клеточной среды, оптимальной для экспрессии генов ВИЧ-1 [7, 8];
- индуцирует ответ на повреждение ДНК, что, как предполагают, может также приводить к остановке клеточного цикла и повышению продукции цитокинов воспаления [6, 7];
- нарушает работу митохондрий, что запускает ряд процессов, которые также могут приводить к апоптозу [9];
- в макрофагах Vpr противодействует специальному клеточному белку, LAMPT5, который транспортирует вирусный белок Env в лизосому [6, 10].

Рис. 2. Активности белка Vpr.
ОТ – обратная транскрипция; Vpr – белок Vpr; Env – белок Env; PIC – прединтеграционный комплекс.
Fig. 2. Activities of Vpr protein.
RT – reverse transcription; Vpr – Vpr protein; Env – Env protein; PIC – pre-integration complex.
Vpr высвобождается продуцирующими клетками и проникает в окружающие В-лимфоциты. В B-лимфоцитах белок Vpr влияет на диверсификацию антител и обладает способностью снижать переключение классов иммуноглобулинов [11]. Отдельное внимание уделяют роли Vpr в усилении вирусной инфекции в неделящихся миелоидных клетках, макрофагах и дендритных клетках, которые позволяют формировать и поддерживать вирусный резервуар, эффективно передавая ВИЧ-1 к CD4+-T-клеткам во время презентации антигена [6, 12].
Белок Vpr рассматривают как один из факторов, способствующих развитию у пациентов нейрокогнитивных ВИЧ-ассоциированных расстройств (HIV-associated neurocognitive disorders, HAND): Vpr может проникать в клетки нервной ткани, является нейротоксином, который индуцирует апоптоз, активирует вирусную репликацию в латентно инфицированных клетках; в нейронах нарушает регуляцию уровней некоторых микроРНК и соответствующих им генов, что также может вызывать нейронную дисфункцию. Более того, ген vpr продолжает экспрессироваться даже при успешной антиретровирусной терапии (АРТ) [13–15].
На протяжении многих лет изучают вопрос корреляции аминокислотных замен в белке Vpr с изменением его функциональных свойств [16, 17]. Определены естественные аминокислотные замены в белке Vpr ВИЧ-1, ассоциированные со степенью развития HAND у лиц, живущих с ВИЧ (ЛЖВ), которые находятся на АРТ [18]. Сравнение генетического разнообразия белка Vpr вариантов ВИЧ-1 у пациентов с быстрым прогрессированием заболевания и у пациентов с долгосрочным отсутствием признаков прогрессирования заболевания в отсутствие АРТ показало, что аминокислотные замены в белке Vpr могут способствовать изменению кинетики репликации вируса и приводить к наблюдаемым различиям в прогрессировании заболевания [19]. При сравнении C-концевой области белка Vpr у вариантов ВИЧ-1 субтипов В и С были определены субтип-специфичные аминокислотные замены, которые могут оказывать влияние на функциональные свойства белка [20].
Практически сразу после описания белка Vpr стали рассматривать возможности создания агентов, ингибирующих репликацию ВИЧ-1 посредством противодействия этому белку: известно большое количество попыток создания антиретровирусных средств как на основе природных, так и синтетических компонентов [17, 21]. Кроме того, белок Vpr содержит эпитопы, распознаваемые T-клетками (https://www.hiv.lanl.gov/content/immunology/maps/helper/Vpr.html, https://www.hiv.lanl.gov/content/immunology/maps/ctl/Vpr.html), и рассматривается в качестве антиген-кандидата для создания анти-ВИЧ-вакцины [22].
Наиболее широко распространенным в России генетическим вариантом ВИЧ-1 на протяжении многих лет остается суб-субтип А6, тогда как в странах Европы, Азии и Америки циркулируют другие варианты вируса [23, 24]. Cо временем сосуществование и взаимодействие ВИЧ-1 суб-субтипа А6 с менее распространенными на территории России генетическими вариантами вируса (субтипом B, циркулирующей рекомбинантной формой CRF02_AG и т.д.) привели к формированию и распространению других рекомбинантов. Недавно проведенные исследования демонстрируют постепенное увеличение с течением времени доли рекомбинантных форм в генетической структуре ВИЧ-1, циркулирующих на территории Российской Федерации, – в частности, за счет CRF63_02A6 [25]. Также в период 2022–2023 гг. на территории России были выявлены две новые формы: CRF133_A6B и CRF157_A6C [26, 27]. Таким образом, несмотря на постепенное изменение состава циркулирующих вариантов ВИЧ-1 в России, молекулярно-эпидемиологический профиль ВИЧ-инфекции по-прежнему сохраняет свою уникальность.
Целью данной работы является изучение особенностей белка Vpr генетических вариантов ВИЧ-1, циркулировавших на территории Московской области в 2019–2020 гг.
Материалы и методы
При выполнении работы были проанализированы клинические образцы цельной крови «наивных» (ранее не получавших лечения) пациентов с ВИЧ-инфекцией ГКУЗ МО «Центр профилактики и борьбы со СПИД» (231 образец). В период 2019–2020 гг. у каждого пациента был проведен однократный забор крови в рамках реализации проекта CARE (https://www.careresearch.eu/, доступ на 01.11.2024). Весь клинический материал был собран и использован в настоящем исследовании с информированного согласия пациентов и на основании одобрения Комитета по биомедицинской этике ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи» Минздрава России (протокол № 16 от 08.02.2019). Параллельно была зарегистрирована и в дальнейшем проанализирована следующая информация о пациентах: пол, возраст, фактор риска инфицирования, дата забора клинического образца, стадия заболевания, показатели вирусной нагрузки (ВН) и иммунный статус пациента (количество CD4+-клеток). В табл. 1 приведены основные характеристики пациентов, включенных в исследование, в зависимости от стадии ВИЧ-инфекции, согласно клиническим рекомендациям Минздрава России1.
Таблица 1. Основные характеристики включенных в исследование ЛЖВ, классифицированных по стадии ВИЧ-инфекции
Table 1. Main characteristics of people living with HIV (PLWH) included in the study, classified by stage of HIV infection
Характеристики Characteristics | 2-я стадия/стадия начальных проявлений Stage 2/stage of initial symptoms | 3-я стадия/ субклиническая стадия Stage 3/subclinical stage | 4-я стадия/стадия вторичных проявлений Stage 4/stage of secondary symptoms |
Всего пациентов, абс. Total number of patients, abs. | 48 | 82 | 101 |
Демографические показатели | Demographics | |||
Мужчины, абс. Males, abs. | 30 | 45 | 71 |
Женщины, абс. Females, abs. | 18 | 37 | 30 |
Возраст, медиана лет, диапазон Age, median age, range | 38 [19; 62] | 38 [21; 70] | 39 [24; 64] |
Путь инфицирования, абс. | Infection route, abs. | |||
Гетеро Hetero | 24 | 59 | 62 |
ПИН IDU | 6 | 11 | 36 |
МСМ MSM | 17 | 9 | 3 |
Неизвестно Unknown | 1 | 3 | 0 |
Лабораторные показатели | Laboratory parameters | |||
CD4, кл/мкл | clones/μL | 599,50 (108–2022) | 474,10 (110–1658) | 229,72 (8–1062) |
Вирусная нагрузка lg РНК, копий/мл Viral load log10 RNA, copies/mL | 5,0 (3,4–7,0) | 4,6 (3,3–6,2) | 5,1 (3,1–6,4) |
Примечание. ПИН – потребители инъекционных наркотиков; МСМ – мужчины, практикующие секс с мужчинами.
Note. IDU – Injecting drug users; MSM – Men having sex with other men.
Выделение провирусной ДНК в составе геномной ДНК осуществляли методом высаливания [28]. Получение продуктов области генома, кодирующей ген vpr, проводили методом гнездовой двухраундовой полимеразной цепной реакции (ПЦР): внешние праймеры – Vif1р (GCAGGTAAGAGAGCAAGCTGAACA) и Vif1о (GTCTCCGCTTCTTCCTGCCATAGGA), внутренние праймеры – Vif2р (GCTaCTCTGGAAAGGTGAAGG) и Vif2о (TACAAGGAGTCTTGGGCTGAC). Полученные ПЦР-продукты были очищены с использованием коммерческого набора для очистки ПЦР-фрагментов – Clean S-Cap («Евроген», Россия), а затем секвенированы дидезокси-методом по Сэнгеру с использованием коммерческого набора BigDye Terminator v. 3.1 Cycle Sequencing Kit (Applied Biosystems, США) и праймеров Vif2р и Vif2о. Сборку и редактирование нуклеотидных последовательностей (н.п.) гена vpr на основе полученных электрофореграмм выполняли с использованием приложения SeqMan II 6.1. (DNASTAR Inc., США).
Предварительное определение генетических вариантов полученных нуклеотидных последовательностей гена vpr ВИЧ-1 осуществляли с применением трех специализированных программ: COMET HIV-1 (https://comet.lih.lu/) [29], REGA HIV-1 Subtyping Tool (Version 3.46) (https://www.genomedetective.com/app/typingtool/hiv) и jpHMM [30]. Затем для уточнения результатов предварительного субтипирования проводили филогенетический анализ методом максимального правдоподобия с использованием программы IQ-TREE [31]. Эталонные последовательности для проведения анализа были выгружены из международной базы данных Лос-Аламосской лаборатории, США (https://www.hiv.lanl, дата обращения 31.10.2024). Попарное и множественное выравнивание исследуемых и эталонных нуклеотидных последовательностей осуществляли с применением модуля ClustalW, интегрированного в программный пакет AliView [32]. Модель замещения нуклеотидов была выбрана при помощи программы jModelTest v. 2.1.7 на основании информационного критерия Акаике (Akaike information criterion, AIC) [33]. Достоверность выведенных филогений оценивали с помощью бутстрэп-теста (bootstrap) и критерия приблизительного отношения правдоподобия Шимодайры–Хасегавы (SH-aLRT) с 1000 послестартовых итераций. Кластеры с поддержкой SH-aLRT > 0,9 считались достоверно установленными. Визуализацию и графическую обработку результатов филогенетического анализа осуществляли в программе iTOL (https://itol.embl.de) [34].
На следующем этапе исследования формировали и анализировали консенсусные последовательности белка Vpr для наиболее распространенных генетических вариантов ВИЧ-1 по результатам исследования. Для этого полученные нуклеотидные последовательности гена vpr были разделены на группы в зависимости от их генетического варианта. Затем нуклеотидные последовательности гена vpr ВИЧ-1 были переведены в аминокислотные с помощью онлайн-инструмента для трансляции, представленного на сайте (https://www.bioinformatics.org/sms2/translate.html). Также для каждого анализируемого генетического варианта ВИЧ-1 на основе полученных аминокислотных последовательностей с помощью инструмента Simple Consensus Maker (https://www.hiv.lanl.gov/content/sequence/CONSENSUS/SimpCon.html) были сформированы общие консенсусные аминокислотные последовательности и сравнены между собой и относительно Vpr_model (последовательность белка Vpr субтипа B, анализируемого при определении его пространственной структуры [2]) с применением программы MEGA v. 10.2.2. При формировании консенсусной последовательности инсерции не учитывались; частота встречаемости, при которой аминокислота (а также стоп-кодон или делеция) в каждой позиции учитывалась в консенсусе, должна была быть больше 50%. При помощи программы IsUnstruct предсказывали расположение неструктурированных участков в консенсусных последовательностях и в Vpr_model [35]. При помощи программы AlphaFold 2 (AlphaFold Protein Structure Database) предсказывали пространственную структуру консенсусных последовательностей и Vpr_model [36].
В программе Chimera предсказанные гексамерные структуры анализируемых последовательностей накладывали друг на друга и на Vpr_model, чтобы определить максимально похожие друг с другом структуры (https://www.rbvi.ucsf.edu/chimera/).
На завершающем этапе исследования изучали вариабельность белка Vpr-A6 (белок Vpr вариантов ВИЧ-1 суб-субтипа А6) у пациентов с разными стадиями заболевания. Для этого полученные аминокислотные последовательности Vpr-A6 были сгруппированы в соответствии со стадией ВИЧ-инфекции пациента, от которого был получен образец. Ранее полученную консенсусную последовательность Vpr-A6 использовали в качестве референсной, относительно нее определяли аминокислотные замены в каждой группе пациентов, с применением программы MEGA v. 10.2.2. (www.megasoftware.net). С применением программного модуля Nonparametric Statistics из пакета Statistica 8.0 (StatSoft Inc., США) выявляли сайты со статистически достоверными различиями в частоте встречаемости у пациентов с разными стадиями заболевания (p < 0,0012 при использовании критерия χ2 с поправкой Бонферрони).
Результаты
При проведении попарного и множественного выравнивания и оценке качества 5 из 231 н.п. были исключены из дальнейшего анализа в связи с низким качеством или длиной нуклеотидных последовательностей (количество вырожденных букв или gap-ов (пробелов) более 1% от общей длины последовательности). В дальнейший анализ были включены 226 н.п., кодирующих ген vpr ВИЧ-1. Все полученные в ходе настоящего исследования нуклеотидные последовательности гена vpr ВИЧ-1 (226) были депонированы в международную базу данных генотипов GenBank со следующими номерами: PV059601–PV059826.
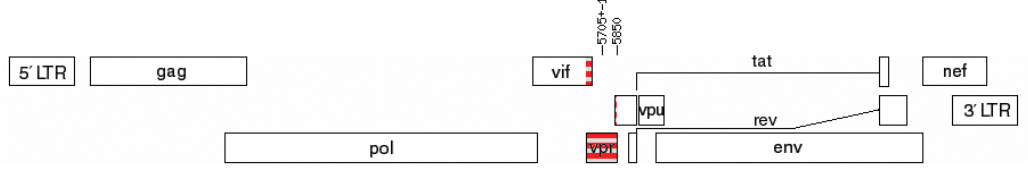
По результатам предварительного субтипирования было установлено, что два образца (0,88%, 2/226), полученные от пациентов со 2-й (1311101072) и с 3-й (1311001115) стадиями заболевания, относились к уникальным рекомбинантам (URFs) ВИЧ-1, образованным фрагментами ВИЧ-1 генетических вариантов B и G. Структуры геномов выявленных URFs_B/G представлены на рис. 3.

Рис. 3. Карта генома с исследуемой областью vpr у образцов 1311001072 (а) и 1311001115 (б).
Fig. 3. Genome map with the studied vpr region in samples 1311001072 (а) и 1311001115 (b).
Данные последовательности были исключены из дальнейшего филогенетического анализа.
По результатам филогенетического анализа 4 (1,77%) н.п. образовали достоверный кластер (SH-aLRT > 0,9) c последовательностями ВИЧ-1 субтипа G, 16 (7,08%) н.п. образовали достоверный кластер (SH-aLRT > 0,9) c последовательностями ВИЧ-1 субтипа B и 11 (4,87%) н.п. были включены в кластер, образованный совместно нуклеотидными последовательностями ВИЧ-1 циркулирующих рекомбинантных форм CRF02_AG и CRF63_02A6 (рис. 4).

Рис. 4. Филогенетический анализ нуклеотидных последовательностей гена vpr ВИЧ-1 (n = 254, модель замещения нуклеотидов – TIM1 + I + G4).
Референсные последовательности выделены красным цветом, исследуемые – черным. Красной звездочкой отмечены нуклеотидные последовательности, отнесенные к потенциальным уникальным рекомбинантам.
Fig. 4. Phylogenetic analysis of nucleotide sequences of the HIV-1 vpr gene (n = 254, nucleotide substitution model – TIM1 + I + G4).
Nucleotide sequences classified as potential unique recombinants are marked with a red asterisk.
Поскольку в исследуемой области генома (vpr) ВИЧ-1 генетические варианты CRF02_AG и CRF63_02A6 обладают максимальным сходством, было принято решение идентифицировать их на основании совместных результатов филогенетического анализа и программы COMET HIV-1 – 10 последовательностей были отнесены к CRF63_02A6. По результатам филогенетического анализа (из-за положения на филогенетическом дереве – опосредованно от остальных) совместно с результатами программы jpHMM (для определения рекомбинантных формы вируса) было принято решение о том, что последовательность 1311001105 (на рисунке отмечена красной звездочкой) может являться потенциальным уникальным рекомбинантом (URF_CRF02/CRF63), структура его генома представлена на рис. 5.

Рис. 5. Карта генома с исследуемой областью vpr у образца 1311001105.
Пунктиром отмечена область, образованная фрагментом ВИЧ-1 рекомбинантных форм CRF02_AG и CRF63_02A6.
Fig. 5. Genome map for the studied vpr region in sample 1311001105.
The dotted line indicates the region formed by the HIV-1 fragment of recombinant forms CRF02_AG and CRF63_02A6.
Последовательность 1311000563 на основе совместных результатов филогенетического анализа и анализа в программе COMET HIV-1 была отнесена к URF_A6/B. Остальные 192 (84,96%) н.п. образовали достоверный кластер с нуклеотидными последовательностями ВИЧ-1 суб-субтипа A6.
Консенсусные последовательности белка Vpr были сформированы для вариантов ВИЧ-1 суб-субтипа А6, субтипа В и рекомбинантной формы CRF63_02A6, выявленных в ходе исследования. Консенсусная аминокислотная последовательность Vpr-А6 была сформирована на основе 192 исследуемых последовательностей, субтипа В – 16, а CRF63_02A6 – 10 соответственно. Данные генетические варианты также являются наиболее распространенными на территории Российской Федерации [24].
В исследуемых аминокислотных последовательностях Vpr-A6 были выявлены инсерции (аминокислотные вставки), делеции (пропуски аминокислот) и стоп-кодоны. Инсерции и делеции наблюдались в аминокислотных последовательностях, полученных от пациентов со всеми стадиями заболевания (табл. 2), тогда как стоп-кодоны – у 6 пациентов с 3-й и 4-й стадиями заболевания.
Таблица 2. Выявленные инсерции и делеции в исследуемых аминокислотных последовательностях Vpr-A6
Table 2. Identified insertions and deletions in the studied amino acid sequences Vpr-A6
Стадия заболевания Infection stage | Инсерции | Insertions | Делеции | Deletions | ||
наименование последовательности sequence name | положение position | наименование последовательности sequence name | положение position | |
2-я стадия Stage 2 | 1311000412 | ins84V85 | 1311000645 | 85 |
1311000512 | ins84I85 | 1311000738 | 85, 86 | |
1311000948 | 85 | |||
3-я стадия Stage 3 | 1311000660 | ins84I85 | 1311000121 | 85, 86 |
1311000278 | 84 | |||
1311000997 | ins84I85 | 1311000601 | 84 | |
1311000613 | 85, 86 | |||
1311000617 | 85, 86 | |||
1311001126 | ins84I85 | 1311000780 | 85, 86 | |
1311001119 | 85 | |||
1311001125 | 85, 86 | |||
4-я стадия Stage 4 | 1311000884 | ins84P85 | 1311000382 | 87, 88 |
1311000599 | 85, 86 | |||
1311000766 | 85 | |||
1311000767 | 85 | |||
1311000919 | 85 | |||
1311001068 | 85, 89 | |||
1311001088 | 85, 86 | |||
1311001089 | 85 | |||
1311001093 | 85 | |||
Все сформированные консенсусные последовательности содержали 96 а.о. (рис. 6).

Рис. 6. Консенсусные последовательности Vpr ВИЧ-1 суб-субтипа А6, субтипа В и рекомбинантной формы CRF63_02A6, выравненные относительно Vpr_model (последовательность белка Vpr, анализируемого при определении пространственной структуры [2]).
Точками обозначены позиции аминокислотных остатков (а.о.), в которых а.о. в консенсусах соответствовали референсу. Аминокислоты классифицированы на основе полярности радикалов. Неполярные аминокислоты: G (глицин), A (аланин), V (валин), L (лейцин), I (изолейцин), P (пролин) отмечены синим цветом; полярные незаряженные аминокислоты: S (серин), T (треонин), C (цистеин), M (метионин), N (аспарагин), Q (глутамин) – зеленым; ароматические аминокислоты: F (фенилаланин), тирозин (Y), W (триптофан), гистидин (H) – желтым; отрицательно заряженные аминокислоты: аспарагиновая кислота (D) и глутаминовая кислота (E) – оранжевым; положительно заряженные аминокислоты: лизин (K), аргинин (R) – красным [37, 38].
Fig. 6. Consensus sequences of Vpr HIV-1 sub-subtype A6, B and CRF63_02A6 genetic variants aligned with the Vpr_model (sequence of the Vpr protein analyzed in determining the spatial structure [2]).
The dots indicate amino acid residues (aa) positions in which the aa in the consensus were the same as in the reference. Non-polar amino acids: G (glycine), A (alanine), V (valine), L (leucine), I (isoleucine), P (proline) – are marked in blue; Polar uncharged amino acids: S (serine), T (threonine), C (cysteine), M (methionine), N (asparagine), Q (glutamine) – green; aromatic amino acids: F (phenylalanine), Tyrosine (Y), W (tryptophan), Histidine (H) – yellow; Polar acidic, negatively charged, amino acids: aspartic acid (D) and glutamic acid (E) – orange; Polar basic, positively charged amino acids: lysine (K), arginine (R) – in red [37, 38].
Первичная структура консенсусных последовательностей суб-субтипа А6, субтипа В и CRF63_02A6 отличалась от Vpr_model в 17, 11 и 15-й позициях соответственно (рис. 6).
В дальнейшем для сравнения пространственных структур при проведении анализа консенсусной последовательности субтипа В анализировали вариант последовательности содержащей в 77-м положении Q, в 84-м – I, при проведении анализа консенсусной последовательности CRF63_02A6 – вариант последовательности, содержащей в 58-м положении R.
На рис. 7 представлены предсказанные профили для неструктурированных участков для Vpr_model, консенсусных последовательностях суб-субтипа А6, субтипа В и CRF63_02A6.

Рис. 7. Сравнение профилей неструктурированных участков для консенсусных последовательностей суб-субтипа А6, субтипа В и CRF63_02A6 и Vpr_model, предсказанные программой IsUnstruct.
a – Vpr_model: развернутые участки с 1–15 и с 86–96 а.о.; б – консенсус суб-субтипа А6: развернутые участки с 1–15 и с 84–96 а.о.; в – консенсус субтипа В: развернутые участки с 1–16 и с 86–96 а.о.; г – консенсус CRF63_02A6: развернутые участки с 1–15 и с 86–96 а.о.
Fig. 7. The comparison of the tertiary structure of the consensus sequences of sub-subtype A6, subtype B and CRF63_02A6 and Vpr_model, predicted by the IsUnstruct program.
a – Vpr_model: unfolded regions from 1 to 15 and from 86 to 96 aa; b – sub-subtype A6 consensus: unfolded regions from 1 to 15 and from 84 to 96 aa; c – subtype B consensus: unfolded regions from 1 to 16 and from 86 to 96 aa; d – CRF63_02A6 consensus: unfolded regions from 1 to 15 and from 86 to 96 aa.
На рис. 8 представлены результаты предсказания пространственной структуры мономерных, димерных, тетрамерных и гексамерных структур, анализируемых последовательностей белка Vpr.

Рис. 8. Мономерные, димерные и олигомерные формы белка Vpr вариантов Vpr_model, суб-субтипа А6, субтипа В, рекомбинантной формы CRF63_02A6, предсказанные программой AlphaFold 2.
a – мономерные формы белка Vpr; б – димерные формы белка Vpr; в – тетрамерная форма белка Vpr; г – гексамерная форма белка Vpr.
Fig. 8. Monomeric, dimeric and oligomeric forms of Vpr protein in Vpr_model, sub-subtype A6, subtype B and CRF63_02A6 variants predicted by the AlphaFold 2 program.
a – monomeric forms of Vpr protein; b – dimeric forms of Vpr protein; c – tetrameric forms of Vpr protein; d – hexameric forms of Vpr protein.
Результаты пространственного выравнивания (сопоставления) предсказанных гексамерных структур представлены на рис. 9.

Рис. 9. Совмещение гексамерных структур Vpr.
Hexamer A6 and Hexamer Vpr_model – гексамер консенсусной последовательности белка суб-субтипа А6 и гексамер Vpr_model; Hexamer B and Hexamer Vpr_model – гексамер консенсусной последовательности белка субтипа B и гексамер Vpr_model; Hexamer CRF63_02A6 and Hexamer Vpr_model – гексамер консенсусной последовательности рекомбинантной формы CRF63_02A6 и гексамер Vpr_model; Hexamer A6 and Hexamer B – гексамер консенсусной последовательности суб-субтипа А6 и гексамер консенсусной последовательности субтипа B; Hexamer A6 and Hexamer CRF63_02A6 – гексамер консенсусной последовательности суб-субтипа А6 и гексамер консенсусной последовательности CRF63_02A6; Hexamer B and Hexamer CRF63_02A6 – гексамер консенсусной последовательности субтипа B и гексамер консенсусной последовательности CRF63_02A6; Среднеквадратичное отклонение между Cα-атомами для разных пар гексамеров показано на рисунке, которое изменяется от 16,9 Å до 37,8 Å.
Fig. 9. Alignment of Vpr hexameric structures.
Hexamer A6 and Hexamer Vpr_model – hexamer of A6 consensus sequence and hexamer Vpr_model; Hexamer B and Hexamer Vpr_model – hexamer of subtype B consensus sequence and hexamer Vpr_model; Hexamer CRF63_02A6 and Hexamer Vpr_model – hexamer of CRF63_02A6 consensus sequence and hexamer Vpr_model; Hexamer A6 and Hexamer B – hexamer of A6 consensus sequence and hexamer of subtype B consensus sequence; Hexamer A6 and Hexamer CRF63_02A6 – hexamer of A6 consensus sequence and hexamer of CRF63_02A6 consensus sequence; Hexamer B and Hexamer CRF63_02A6 – hexamer of subtype B consensus sequence and hexamer of CRF63_02A6 consensus sequence; The root mean square deviation between Cα atoms for different pairs of hexamers is shown in the figure, which varies from 16.9 Å to 37.8 Å.
При оценке вариабельности Vpr-А6 у пациентов с разными стадиями ВИЧ-инфекции было выявлено 14 замен, имеющих статистически значимые различия (p < 0,05) в частоте их встречаемости (табл. 3).
Таблица 3. Аминокислотные замены Vpr-A6 со статистически значимыми различиями по частоте встречаемости в группах ЛЖВ с разными стадиями заболевания
Table 3. Vpr-A6 amino acid substitutions with statistically significant differences in frequency of occurrence in groups of PLWH with different stages of the disease
Позиция Position | Мутация Mutation | Стадия 2 Stage 2 | Стадия 3 Stage 3 | Стадия 4 Stage 4 | p2‒3 | p2‒4 | p3–4 |
13 | E13A | 2 | 0 | 0 | 0,0465 | 0,0190 | – |
15 | Y15H | 6 | 8 | 23 | – | – | 0,0444 |
15 | Y15F | 1 | 5 | 1 | – | – | 0,0353 |
19 | M19V | 3 | 0 | 2 | 0,0143 | – | – |
20 | L20I | 0 | 3 | 0 | – | – | 0,0390 |
45 | H45Q | 6 | 6 | 3 | – | 0,0054 | – |
55 | E55V | 2 | 0 | 0 | 0,0465 | 0,0190 | – |
61 | I61T | 6 | 2 | 8 | 0,0107 | – | – |
72 | F72Y | 2 | 0 | 0 | 0,0465 | 0,0190 | – |
77 | Q77H | 5 | 4 | 4 | – | 0,0451 | – |
85 | Q85H | 1 | 0 | 7 | – | – | 0,0219 |
87 | R87S | 5 | 3 | 3 | – | 0,0194 | – |
93 | S93T | 0 | 3 | 0 | – | – | 0,0390 |
94 | S94N | 1 | 3 | 0 | – | – | 0,0390 |
Примечание. Значения p-value представлены для позиций с p < 0,05; позиции с p ≥ 0,05 отмечены знаком «–». Достоверно значимыми считали различия с p-value с поправкой Бонферрони (p < 0,0012).
Note. The p-values are presented for items with p < 0.05; items with p ≥ 0.05 are marked with ‘–’. Differences with p-value with Bonferroni correction (p < 0.0012)
C учетом поправки Бонферрони (p < 0,0012) не было выявлено ни одного сайта со статистически значимыми различиями в частоте встречаемости у пациентов с разными стадиями заболевания.
Обсуждение
В настоящее время во всем мире продолжается рост генетического разнообразия ВИЧ-1, которое является одним из препятствий для разработки эффективных средств профилактики и терапии ВИЧ-инфекции [39]. Более того, результаты исследований позволяют предположить, что разные генетические варианты ВИЧ-1 способны обуславливать различные клинические проявления и скорость прогрессирования заболевания, а также оказывать влияние на эффективность лечения [40]. Регулярно проводятся работы, посвященные изучению степени влияния отдельных вирусных белков на течение ВИЧ-инфекции [14, 41–43]. Также изучают аминокислотные замены в вирусных белках, которые могут оказывать влияние на прогрессирование ВИЧ-инфекции, отдельное внимание уделяется характерным субтип-специфичным аминокислотным заменам [18, 44, 45]. В ранее проведенных исследованиях генетического разнообразия белка Vpr наиболее широко распространенного в России суб-субтипа А6 отмечали его низкий уровень изменчивости и, в связи с этим, определяли белок Vpr как перспективную мишень для разработки средств терапии [46]. Также для белка Vpr суб-субтипа А6 вариантов вируса, циркулирующих в разных регионах России, предварительно не было отмечено наличия характерных особенностей [47]. Настоящее исследование направлено на изучение особенностей белка Vpr наиболее широко распространенных в России генетических вариантов ВИЧ-1 на примере вариантов вирусов, циркулировавших в Московской области в 2019–2020 гг., и сравнение генетической вариабельности белка Vpr-A6 у пациентов с разными стадиями заболевания.
По результатам исследования было установлено, что большинство (84,96%) нуклеотидных последовательностей vpr относились к ВИЧ-1 суб-субтипа А6, вторым по распространенности стал субтип B (7,08%), затем – рекомбинантная форма CRF63_02A6 (4,87%), что согласуется с результатами исследования генетического разнообразия ВИЧ-1 на территории Российской Федерации [24]. Две последовательности были идентифицированы как уникальные B/G-рекомбинантные формы, что также согласуется с ранее представленными данными о выявлении в России уникальных B/G-рекомбинантных форм [48]. Одна последовательность гена vpr была определена как уникальная A6/B-рекомбинантная форма, что подкрепляется данными о формировании различных рекомбинантных форм между ВИЧ-1 суб-субтипа А6 и субтипа В на территории России [49].
Шесть из 192 (3,13%) последовательностей Vpr-A6, полученные от пациентов с разными стадиями ВИЧ-инфекции, содержали инсерцию между 84-м и 85-м положениями аминокислотных остатков. Двадцать из 192 (10,42%) последовательностей Vpr-A6, полученные от пациентов с разными стадиями заболевания, содержали делеции в 84–89-м положениях, при этом 50% данных последовательностей содержали сразу две делеции. Наиболее часто делеции встречались в 85-м (85%, 17/20) и в 86-м (40%, 8/20) положениях (табл. 2). Ранее уже отмечалось наличие делеций в 85, 86 и 89-й позициях в белке Vpr вариантов суб-субтипа А6 [46]. Преждевременные стоп-кодоны были выявлены у 6 пациентов с 3-й и 4-й стадиями заболевания. Известны исследования закономерностей и частоты встречаемости стоп-кодонов в участках провирусной ДНК, кодирующих протеазу, обратную транскриптазу и интегразу [50, 51]. Однако аналогичных работ по изучению распространенности стоп-кодонов в провирусной ДНК, кодирующей белок Vpr, пока не проводилось. В целом в настоящее время существует предположение, что дефектные провирусы могут обладать биологической активностью: образующиеся на их основе транскрипты и соответствующие им белки могут участвовать в стимуляции иммунного ответа, последующей хронической активации иммунной системы и быть серьезным препятствием для разработки средств эрадикации ВИЧ [52].
Консенсусные последовательности анализируемых вариантов ВИЧ-1 в нескольких позициях, предположительно участвующих в формировании пространственной структуры белка, содержали замены относительно референсной последовательности – Vpr_model: замену D17E – суб-субтип А6, субтип В, CRF63_02A6; T19M – суб-субтип А6, T55E – суб-субтип А6, CRF63_02A6; T55A – субтип В; L60I – суб-субтип А6, субтип В, CRF63_02A6 и R77Q – суб-субтип А6, субтип В, CRF63_02A6. При этом замены T55E, T55A и R77Q приводили к изменению химических свойств аминокислотного остатка в заданном положении.
Предсказание пространственных структур консенсусных последовательностей и референсной последовательности Vpr_model определило, что в анализируемых последовательностях структурированные участки белка преимущественно приходились на область с 16-го по 85-й а.о., что совпадало с областью, в которой картировались эпитопы в белке Vpr (рис. 1, 7).
Предсказание олигомерных структур белка Vpr консенсусных последовательностей и Vpr_model продемонстрировало различия среди тетрамерных и гексамерных форм. При пространственном выравнивании гексамерных форм было определено, что наибольшее среднеквадратичное отклонение между Сα-атомами (RMSD) составило 37,8 Å для пары гексамера консенсусной последовательности субтипа B и гексамера консенсусной последовательности CRF63_02A6, а наименьшее – 16,9 Å для пары гексамера консенсусной последовательности рекомбинантной формы CRF63_02A6 и гексамера Vpr_model (рис. 9). RMSD является количественной мерой сходства между двумя белковыми структурами, и наименьшее значение RMSD между олигомерными формами указывает на их структурное сходство.
Таким образом, имеющиеся особенности белка Vpr у различных вариантов ВИЧ-1 могут повлиять на формирование олигомерных форм белка. Учитывая высокую значимость процесса олигомеризации, который влияет на включение белка Vpr в вирионы и, как следствие, определяет возможность участия белка Vpr в ранних стадиях репликации вируса, можно утверждать, что существующие особенности могут повлиять на функциональные свойства белка Vpr [4].
В ранее выполненном исследовании динамики вариабельности гена vpr у пациентов, инфицированных ВИЧ-1 субтипа С, отмечалось постепенное увеличение генетического разнообразия vpr в популяции вируса у пациента в течение первого года развития заболевания [53]. Однако сравнение белка Vpr у вариантов вируса, выделенных от пациентов с разными стадиями заболевания в Китае, не выявило существенных различий аминокислот в функционально значимых областях [54]. В данном исследовании также не было обнаружено аминокислотных замен в белке Vpr-A6, имеющих статистически значимые различия в частоте встречаемости у пациентов с разными стадиями заболевания, что подтверждает ранее сделанные выводы о низкой вариабельности белка Vpr-A6 и перспективности его использования для создания средств терапии ВИЧ-инфекции [46, 47].
Ограничениями проведенного исследования является относительно небольшая выборка, в том числе не-A6-вариантов, а также изучение вариантов вируса, циркулирующих в пределах одного региона – Московской области.
Заключение
Впервые проведено сравнение особенностей белков Vpr генетических вариантов ВИЧ-1, наиболее широко распространенных на территории Российской Федерации (A6, B, CRF63). Было установлено, что присутствующие особенности могут повлиять на формирование олигомерных форм белка. Принимая во внимание важность процесса олигомеризации белка Vpr, можно предположить, что имеющиеся различия могут приводить к разной функциональной активности белка Vpr у вариантов ВИЧ-1. Вместе с тем сравнение генетического разнообразия Vpr-A6 у пациентов с разными стадиями ВИЧ-инфекции не выявило достоверно значимых аминокислотных замен, что подтверждает данные о низкой вариабельности белка Vpr внутри вариантов ВИЧ-1 суб-субтипа А6 и о возможности его использования для разработки средств терапии ВИЧ-инфекции.
1 Минздрав РФ. Клинические рекомендации. ВИЧ-инфекция у взрослых; 2024. Available at: https://cr.minzdrav.gov.ru/schema/79_2
Об авторах
Анна Игоревна Кузнецова
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
Автор, ответственный за переписку.
Email: a-myznikova@list.ru
ORCID iD: 0000-0001-5299-3081
канд. биол. наук, заведующая лабораторией вирусов лейкозов, ведущий научный сотрудник Института вирусологии им. Д.И. Ивановского
Россия, 123098, г. МоскваАнастасия Александровна Антонова
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
Email: aantonova1792@gmail.com
ORCID iD: 0000-0002-9180-9846
канд. биол. наук, научный сотрудник лаборатории вирусов лейкозов, Институт вирусологии им. Д.И. Ивановского
Россия, 123098, г. МоскваЕкатерина Александровна Макеева
ФГАОУ ВО «Московский политехнический университет»
Email: makeevakaty13@gmail.com
ORCID iD: 0009-0005-7085-3361
студентка факультета химической технологии и биотехнологии
Россия, 107023, г. МоскваКристина Вячеславовна Ким
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
Email: kimsya99@gmail.com
ORCID iD: 0000-0002-4150-2280
младший научный сотрудник лаборатории вирусов лейкозов, Институт вирусологии им. Д.И. Ивановского
Россия, 123098, г. МоскваЯна Михайловна Мунчак
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
Email: yana_munchak@mail.ru
ORCID iD: 0000-0002-4792-8928
младший научный сотрудник лаборатории вирусов лейкозов, Институт вирусологии им. Д.И. Ивановского
Россия, 123098, г. МоскваЕкатерина Никитична Меженская
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
Email: belokopytova.01@mail.ru
ORCID iD: 0000-0002-3110-0843
канд. биол. наук, научный сотрудник лаборатории вирусов лейкозов, Институт вирусологии им. Д.И. Ивановского
Россия, 123098, г. МоскваЕлена Александровна Орлова-Морозова
ГБУЗ МО «Центр профилактики и борьбы со СПИД»
Email: orlovamorozova@gmail.com
ORCID iD: 0000-0003-2495-6501
канд. мед. наук, заведующая амбулаторно-поликлиническим отделением
Россия, 140053, г. Котельники, Московская областьАлександр Юрьевич Пронин
ГБУЗ МО «Центр профилактики и борьбы со СПИД»
Email: alexanderp909@gmail.com
ORCID iD: 0000-0001-9268-4929
канд. мед. наук, главный врач
Россия, 140053, г. Котельники, Московская областьАлексей Геннадьевич Прилипов
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России
Email: a_prilipov@mail.ru
ORCID iD: 0000-0001-8755-1419
д-р биол. наук, ведущий научный сотрудник, заведующий лабораторией молекулярной генетики Института вирусологии им. Д.И. Ивановского
Россия, 123098, г. МоскваОксана Валериановна Галзитская
ФГБУ «Национальный исследовательский центр эпидемиологии и микробиологии имени Н.Ф. Гамалеи» Минздрава России; ФБУН «Институт теоретической и экспериментальной биофизики» Российской академии наук
Email: ogalzit@vega.protres.ru
ORCID iD: 0000-0002-3962-1520
д-р физ.-мат. наук, заведующая лабораторией биоинформатики, главный научный сотрудник ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи», Институт вирусологии им. Д.И. Ивановского
Россия, 123098, г. Москва; 142290, г. Пущино, Московская областьСписок литературы
- Kogan M., Rappaport J. HIV-1 accessory protein Vpr: Relevance in the pathogenesis of HIV and potential for therapeutic intervention. Retrovirology. 2011; 8: 25. https://doi.org/10.1186/1742-4690-8-25
- Morellet N., Bouaziz S., Petitjean P., Roques B.P. NMR structure of the HIV-1 regulatory protein VPR. J. Mol. Biol. 2003; 327(1): 215–27. https://doi.org/10.1016/s0022-2836(03)00060-3
- Sawaya B.E., Khalili K., Gordon J., Taube R., Amini S. Cooperative interaction between HIV-1 regulatory proteins Tat and Vpr modulates transcription of the viral genome. J. Biol. Chem. 2000; 275(45): 35209–14. https://doi.org/10.1074/jbc.M005197200
- Fritz J.V., Dujardin D., Godet J., Didier P., De Mey J., Darlix J.L., et al. HIV-1 Vpr oligomerization but not that of Gag directs the interaction between VPR and GAG. J. Virol. 2010; 84(3): 1585–96. https://doi.org/10.1128/JVI.01691-09
- Venkatachari N.J., Walker L.A., Tastan O., Le T., Dempsey T.M., Li Y., et al. Human immunodeficiency virus type 1 Vpr: oligomerization is an essential feature for its incorporation into virus particles. Virol. J. 2010; 7: 119. https://doi.org/10.1186/1743-422X-7-119
- Nodder S.B., Gummuluru S. Illuminating the role of VPR in HIV infection of myeloid cells. Front. Immunol. 2019; 10: 1606. https://doi.org/10.3389/fimmu.2019.01606
- Vanegas-Torres C.A., Schindler M. HIV-1 Vpr functions in primary CD4+ T Cells. Viruses. 2024; 16(3): 420. https://doi.org/10.3390/ v16030420
- Zhang F., Bieniasz P.D. HIV-1 VPR induces cell cycle arrest and enhances viral gene expression by depleting CCDC137. Elife. 2020; 9: e55806. https://doi.org/10.7554/eLife.55806
- Huang C.Y., Chiang S.F., Lin T.Y., Chiou S.H., Chow K.C. HIV-1 VPR triggers mitochondrial destruction by impairing Mfn2-mediated ER-mitochondria interaction. PLoS One. 2012; 7(3): e33657. https://doi.org/10.1371/journal.pone.0033657
- Zhao L., Wang S., Xu M., He Y., Zhang X., Xiong Y., et al. VPR counteracts the restriction of LAPTM5 to promote HIV-1 infection in macrophages. Nat. Commun. 2021; 12(1): 3691. https://doi.org/10.1038/s41467-021-24087-8
- Eldin P., Péron S., Galashevskaya A., Denis-Lagache N., Cogné M., Slupphaug G., et al. Impact of HIV-1 Vpr manipulation of the DNA repair enzyme UNG2 on B lymphocyte class switch recombination. J. Transl. Med. 2020; 18(1): 310. https://doi.org/10.1186/s12967-020-02478-7
- Casey Klockow L., Sharifi H.J., Wen X., Flagg M., Furuya A.K., Nekorchuk M., et al. The HIV-1 protein Vpr targets the endoribonuclease Dicer for proteasomal degradation to boost macrophage infection. Virology. 2013; 444(1-2): 191–202. https://doi.org/10.1016/j.virol.2013.06.010
- Li G., Makar T., Gerzanich V., Kalakonda S., Ivanova S., Pereira E.F.R., et al. HIV-1 VPR-induced proinflammatory response and apoptosis are mediated through the Sur1-Trpm4 channel in astrocytes. mBio. 2020; 11(6): e02939–20. https://doi.org/10.1128/mbio.02939-20
- Mukerjee R., Chang J.R., Del Valle L., Bagashev A., Gayed M.M., Lyde R.B., et al. Deregulation of microRNAs by HIV-1 VPR protein leads to the development of neurocognitive disorders. J. Biol. Chem. 2011; 286(40): 34976–85. https://doi.org/10.1074/jbc.M111.241547
- James T., Nonnemacher M.R., Wigdahl B., Krebs F.C. Defining the roles for VPR in HIV-1-associated neuropathogenesis. J. Neurovirol. 2016; 22(4): 403–15. https://doi.org/10.1007/s13365-016-0436-5
- Fabryova H., Strebel K. VPR and its cellular interaction partners: R we there yet? Cells. 2019; 8(11): 1310. https://doi.org/10.3390/cells8111310
- González M.E. The HIV-1 VPR protein: A multifaceted target for therapeutic intervention. Int. J. Mol. Sci. 2017; 18(1): 126. https://doi.org/10.3390/ijms18010126
- Dampier W., Antell G.C., Aiamkitsumrit B., Nonnemacher M.R., Jacobson J.M., Pirrone V., et al. Specific amino acids in HIV-1 Vpr are significantly associated with differences in patient neurocognitive status. J. Neurovirol. 2017; 23(1): 113–24. https://doi.org/10.1007/s13365-016-0462-3
- Hadi K., Walker L.A., Guha D., Murali R., Watkins S.C., Tarwater P., et al. Human immunodeficiency virus type 1 VPR polymorphisms associated with progressor and non-progressor individuals alter VPR-associated functions. J. Gen. Virol. 2014; 95(3): 700–11. https://doi.org/10.1099/vir.0.059576-0
- Colle J.H., Rose T., Rouzioux Ch., Garcia A. Two highly variable Vpr84 and Vpr85 residues within the HIV-1-Vpr C-terminal protein transduction domain control transductionnal activity and define a clade specific polymorphism. World Journal of AIDS. 2014; (4): 148–55. https://doi.org/10.4236/wja.2014.4201
- Hagiwara K., Ishii H., Murakami T., Takeshima S.N., Chutiwitoonchai N., Kodama E.N., et al. Synthesis of a VPR-binding derivative for use as a novel HIV-1 inhibitor. PLoS One. 2015; 10(12): e0145573. https://doi.org/10.1371/journal.pone.0145573
- Milani A., Baesi K., Agi E., Marouf G., Ahmadi M., Bolhassani A. HIV-1 accessory proteins: which one is potentially effective in diagnosis and vaccine development? Protein Pept. Lett. 2021; 28(6): 687–98. https://doi.org/10.2174/0929866528999201231213610
- Bbosa N., Kaleebu P., Ssemwanga D. HIV subtype diversity worldwide. Curr. Opin. HIV AIDS. 2019; 14(3): 153–60. https://doi.org/10.1097/COH.0000000000000534
- Антонова А.А., Кузнецова А.И., Ожмегова Е.Н., Лебедев А.В., Казеннова Е.В., Ким К.В. и др. Генетическое разнообразие ВИЧ-1 на современном этапе эпидемии в Российской Федерации: увеличение распространенности рекомбинантных форм. ВИЧ-инфекция и иммуносупрессии. 2023; 15(3): 61–72. https://doi.org/10.22328/2077-9828-2023-15-3-61-72 https://elibrary.ru/tpwttn
- Antonova A., Kazennova E., Lebedev A., Ozhmegova E., Kuznetsova A., Tumanov A., et al. Recombinant forms of HIV-1 in the last decade of the epidemic in the Russian Federation. Viruses. 2023; 15(12): 2312. https://doi.org/10.3390/v15122312
- Maksimenko L.V., Sivay M.V., Totmenin A.V., Shvalov A.N., Skudarnov S.E., Ostapova T.S., et al. Novel HIV-1 A6/B recombinant forms (CRF133_A6B and URF_A6/B) circulating in Krasnoyarsk region, Russia. J. Infect. 2022; 85(6): 702–69. https://doi.org/10.1016/j.jinf.2022.10.001
- Halikov M.R., Ekushov V.E., Totmenin A.V., Gashnikova N.M., Antonets M.E., Tregubchak T.V., et al. Identification of a novel HIV-1 circulating recombinant form CRF157_A6C in Primorsky Territory, Russia. J. Infect. 2024; 88(2): 180–2. https://doi.org/10.1016/j.jinf.2023.11.005
- Miller S.A., Dykes D.D., Polesky H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic. Acids. Res. 1988; 16(3): 1215. https://doi.org/10.1093/nar/16.3.1
- Struck D., Lawyer G., Ternes A.M., Schmit J.C., Bercoff D.P. COMET: adaptive context-based modeling for ultrafast HIV-1 subtype identification. Nucleic Acids Res. 2014; 42(18): e144. https://doi.org/10.1093/nar/gku739
- Schultz A.K., Bulla I., Abdou-Chekaraou M., Gordien E., Morgenstern B., Zoaulim F., et al. jpHMM: recombination analysis in viruses with circular genomes such as the hepatitis B virus. Nucleic Acids Res. 2012; 40(Web Server issue): W193–8. https://doi.org/10.1093/nar/gks414
- Nguyen L.T., Schmidt H.A., von Haeseler A., Minh B.Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015; 32(1): 268–74. https://doi.org/10.1093/molbev/msu300
- Larsson A. AliView: a fast and lightweight alignment viewer and editor for large datasets. Bioinformatics. 2014; 30(22): 3276–8. https://doi.org/10.1093/bioinformatics/btu531
- Darriba D., Taboada G.L., Doallo R., Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods. 2012; 9(8): 772. https://doi.org/10.1038/nmeth.2109
- Letunic I., Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021; 49(W1): W293–6. https://doi.org/10.1093/nar/gkab301
- Lobanov M.Y., Sokolovskiy I.V., Galzitskaya O.V. IsUnstruct: prediction of the residue status to be ordered or disordered in the protein chain by a method based on the Ising model. J. Biomol. Struct. Dyn. 2013; 31(10): 1034–43. https://doi.org/10.1080/07391102.2012.718529
- Jumper J., Evans R., Pritzel A., Green T., Figurnov M., Ronneberger O., et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021; 596(7873): 583–9. https://doi.org/10.1038/s41586-021-03819-2
- Березов Т.Т., Коровкин Б.Ф. Биологическая химия. М.: Медицина; 1998.
- Lobanov M.Y., Pereyaslavets L.B., Likhachev I.V., Matkarimov B.T., Galzitskaya O.V. Is there an advantageous arrangement of aromatic residues in proteins? Statistical analysis of aromatic interactions in globular proteins. Comput. Struct. Biotechnol. J. 2021; 19: 5960–8. https://doi.org/10.1016/j.csbj.2021.10.036
- Nair M., Gettins L., Fuller M., Kirtley S., Hemelaar J. Global and regional genetic diversity of HIV-1 in 2010-21: systematic review and analysis of prevalence. Lancet Microbe. 2024; 5(11): 100912. https://doi.org/10.1016/S2666-5247(24)00151-4
- Bouman J.A., Venner C.M., Walker C., Arts E.J., Regoes R.R. Per-pathogen virulence of HIV-1 subtypes A, C and D. Proc. Biol. Sci. 2023; 290(1998): 20222572. https://doi.org/10.1098/rspb.2022.2572
- Sami Saribas A., Cicalese S., Ahooyi T.M., Khalili K., Amini S., Sariyer I.K. HIV-1 Nef is released in extracellular vesicles derived from astrocytes: evidence for Nef-mediated neurotoxicity. Cell Death Dis. 2017; 8(1): e2542. https://doi.org/10.1038/cddis.2016.467
- Cafaro A., Schietroma I., Sernicola L., Belli R., Campagna M., Mancini F., et al. Role of HIV-1 tat protein interactions with host receptors in HIV infection and pathogenesis. Int. J. Mol. Sci. 2024; 25(3): 1704. https://doi.org/10.3390/ijms25031704
- Khan N., Geiger J.D. Role of Viral Protein U (VPU) in HIV-1 infection and pathogenesis. Viruses. 2021; 13(8): 1466. https://doi.org/10.3390/v13081466
- Ruiz A.P., Ajasin D.O., Ramasamy S., DesMarais V., Eugenin E.A., Prasad V.R. A naturally occurring polymorphism in the HIV-1 tat basic domain inhibits uptake by bystander cells and leads to reduced neuroinflammation. Sci. Rep. 2019; 9(1): 3308. https://doi.org/10.1038/s41598-019-39531-5
- Lebedev A., Kim K., Ozhmegova E., Antonova A., Kazennova E., Tumanov A., et al. Rev protein diversity in HIV-1 group M clades. Viruses. 2024; 16(5): 759. https://doi.org/10.3390/v16050759
- Лапавок И.А. Анализ полиморфизма неструктурных областей генома варианта ВИЧ-1, доминирующего в России: Автореф. дисс. … канд. биол. наук. М.; 2009. https://elibrary.ru/nkranl
- Антонова А.А., Лебедев А.В., Ожмегова Е.Н., Шлыкова А.В., Лаповок И.А., Кузнецова А.И. Вариабельность неструктурных белков у вариантов ВИЧ-1 суб-субтипа А6 (Retroviridae: Orthoretrovirinae: Lentivirus: Human immunodeficiency virus-1, sub-subtype A6), циркулирующих в разных регионах Российской Федерации. Вопросы вирусологии. 2024; 69(5): 470–80. https://doi.org/10.36233/0507-4088-262 https://elibrary.ru/wbbkuq
- Murzakova A., Kireev D., Baryshev P., Lopatukhin A., Serova E., Shemshura A., et al. Molecular epidemiology of HIV-1 subtype G in the Russian Federation. Viruses. 2019; 11(4): 348. https://doi.org/10.3390/v11040348
- Щемелев А.Н., Семенов А.В., Останкова Ю.В., Найденова Е.В., Зуева Е.Б., Валутите Д.Э. и др. Генетическое разнообразие вируса иммунодефицита человека (ВИЧ-1) в Калининградской области. Вопросы вирусологии. 2022; 67(4): 310–21. https://elibrary.ru/bkswno
- Makinson A., Masquelier B., Taieb A., Peytavin G., Waldner-Combernoux A., Collin G., et al. Presence of numerous stop codons in HIV-1 reverse transcriptase proviral DNA sequences from patients with virological response to HAART. AIDS. 2006; 20(9): 1327–9. https://doi.org/10.1097/01.aids.0000232242.51286.7b
- Alidjinou E.K., Deldalle J., Robineau O., Hallaert C., Meybeck A., Huleux T., et al. Routine drug resistance testing in proviral HIV-1 DNA: Prevalence of stop codons and hypermutation, and associated factors. J. Med. Virol. 2019; 91(9): 1684–7. https://doi.org/10.1002/jmv.25474
- Бобкова М.Р. Дефектные провирусы ВИЧ: возможное участие в патогенезе ВИЧ-инфекции. Вопросы вирусологии. 2024; 69(5): 399–414. https://doi.org/10.36233/0507-4088-261 https://elibrary.ru/pselci
- Rossenkhan R., Novitsky V., Sebunya T.K., Musonda R., Gashe B.A., Essex M. Viral diversity and diversification of major non-structural genes vif, vpr, vpu, tat exon 1 and rev exon 1 during primary HIV-1 subtype C infection. PLoS One. 2012; 7(5): e35491. https://doi.org/10.1371/journal.pone.0035491
- Shen C., Gupta P., Wu H., Chen X., Huang X., Zhou Y., et al. Molecular characterization of the HIV type 1 vpr gene in infected Chinese former blood/plasma donors at different stages of diseases. AIDS Res. Hum. Retroviruses. 2008; 24(4): 661–6. https://doi.org/10.1089/aid.2007.0270
Дополнительные файлы

